Metabolism and Physiological Effects of Cyclic Fatty Acids Formed from Linoleic and alpha-Linolenic Acids during Frying
The Author: Jean Louis Sébédio, UMR 1019, Unité de Nutrition Humaine, Plateforme d’exploration du métabolisme, INRA centre de Theix, 63122 St Genes Champanelle, France
1. Introduction
Fatty acids of vegetable oils subjected to the high temperatures during refining (deodorization process) and during frying can undergo cyclization. While saturated and oleic acids are hardly affected, α-linolenic acid is about ten times more sensitive than linoleic acid. Structures of the major cyclic fatty acid isomers (CFAM) formed from linoleic and α-linolenic acids have been elucidated using a combination of mass spectrometry, gas-liquid chromatography and infrared spectroscopy (see the web page on analysis of cyclic fatty acids)[1].
Figure 1. Basic structures of the cyclic fatty acids formed from α-linolenate.
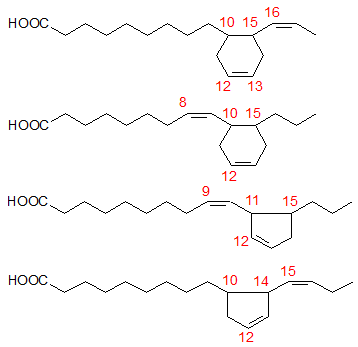
CFAM formed from linolenic acid are basically a mixture of dienoic di-substituted 5- and 6-carbon-membered ring isomers, while those arising from linoleic acid are mainly 5-carbon-membered ring monounsaturated isomers (see the web page on the chemistry of cyclization). Many nutritional studies have been carried out either with heated oils containing many different compounds (polymers, oxidized compounds, CFAM, trans fatty acid isomers, and so forth) or partially enriched fractions of cyclic fatty acids. Few reports have dealt with pure synthesised or isolated CFAM fractions.
Figure 2. Some of the fatty acids produced during cyclization of linoleate.
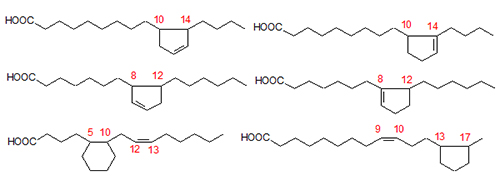
This review only deals with studies using pure characterized cyclic monomer fractions. In order to isolate and purify CFAM fractions, methods were developed to prepare CFAMs from heated oils. These are a combination of column chromatography, urea inclusion, and preparative high-performance liquid chromatography (HPLC) [1,2].
2. Metabolism and Incorporation into Biological Tissues
Earlier studies showed that CFAM were absorbed efficiently in the intestines [3]. More recently, Martin et al. [4] demonstrated that the effects of CFAM upon intestinal metabolism are greatly influenced by their positioning within the triacylglycerol and that the structure of CFAM influences their lymphatic recovery only when they are absorbed as free fatty acids. A selective binding to the Fatty Acid Binding Protein (FABP) was suggested to explain these results.
Studies using hepatic subcellular fractions showed a lower oxygen consumption and a lower activity of carnitine palmitoyl transferase compared to 16:0 and 18:2n-6 for mitochondrial oxidation [5]. For peroxisomal oxidation, CFAM formed from linoleic acid showed the same kinetic parameters as 18:2n-6 and 16:0 (control), while a lower acyl-CoA oxidase activity for CFAM isolated from 18:3 than for the other fatty acids was observed. Both types of CFAMs were poor substrates for acyl-CoA synthase and consequently, the activation of these CFAMs into their CoA derivatives may be the limiting step in their metabolism.
A study in vivo using rats force-fed with 1-14C-labeled CFAMs of both types showed that these unusual fatty acids can be used for energy production [9]. Furthermore, no significant differences in CO2 production were found 24 h after the administration of CFAM and their respective precursors (linoleic and α-linolenic acids). These data demonstrate that at least for the first beta oxidation cycle CFAM are oxidized in a similar way as both essential fatty acids. For the CFAM isolated from 18:2, the larger part of the radioactivity recovered in the lower portion of the gastrointestinal tract and in the urine would suggest that these cyclic fatty acids might be poorly metabolized compared to their precursor, linoleic acid [6].
CFAM can be incorporated into all types of animal tissues, both in non-phosphorus lipids (NL) and in phospholipids (PL), but the incorporation into tissue lipids may be selective (type of lipid classes and type of tissues). Furthermore, for adult rats, incorporation of CFAM formed from α-linolenic acid is twice as much as that of CFAM formed from linoleic acid in heart and liver PL. The CFAM composition of adipose tissue reflected that of the original CFAM mixtures fed to the animal, while differences existed between CFAM fed those found in PL and NL fractions of tissues, with a relative increase of components having a 5-carbon-membered ring [7].
The size of the ring is not the only factor influencing incorporation of CFAM. For example, cis vs. trans configuration at the ring substitution, propyl vs. butyl substitution on the ring, and double bond position and geometry also contribute to the observed differences.
Similar results were reported by Ribot et al. when looking at incorporation of CFAM isolated from heated linseed oil in rat heart cell cultures [8]. Currently, it is very difficult to explain these selective incorporations as in these studies all CFAM analyses were carried out after hydrogenation of the total fatty acid methyl esters. Consequently the complexity of the CFAM mixtures was reduced. Methods enabling isolation and quantification of CFAM without hydrogenation must be utilized in order to better understand these selective incorporations of CFAM into biological tissues.
3. Biological Effects
Many studies were done with heated oils containing cyclic monomers, but in most cases physiological effects observed could not be related specifically to the presence of these fatty acids. In this chapter, we outline the major studies carried out with isolated CFAM fractions, but more detailed information is available in a recently published review [7].
Evidence that CFAM could be harmful was presented by Crampton et al. in the 1950s [9,10], but the high quantities of cyclic monomers in the diets constituted the weak point of the work. Some early studies were also performed by Iwaoka and Perkins [11], who prepared CFAM fractions by cyclization of methyl linolenate at high temperatures under nitrogen. These cyclic monomers were given at levels ranging from 75 to 1,500 ppm in diets with 15% protein until rats reached 200 g. They were then incubated with 0.2 mL uniformly labelled cyclic monomers. Control rats received the same quantity of labelled linoleate. Only a low quantity of radioactivity was detected as CO2 during 48 h (14% compared to 36% in controls). Urine contained 40% of the radioactivity compared to 1% in controls. The amount of cyclic monomers previously ingested had no effect on the percentages of radioactivity detected in urine and CO2.
In this study, the increase of radioactivity of urine in rats suggests that a detoxification mechanism exists, and this is quite consistent with the preferential incorporation of 5-carbon-membered ring cyclic monomers in tissue lipids observed in many experiments. The very low toxicity of 6-carbon-membered ring cyclic monomers, as indicated by Iwaoka and Perkins, is explained by their rapid excretion. This point is actually only hypothetical and should be further studied by comparing synthetic 5- and 6-carbon-membered ring cyclic labeled fatty acids in order to look at their excretion in urine. Some previous results [12] demonstrated that compounds resulting from detoxification, such as glucuronides, appeared in urine of rats after ingestion of heated oil containing cyclic monomers. Furthermore, an induction of hepatic drug-metabolizing enzymes was observed in rats after ingestion of isolated CFAM formed from linolenic acid [13]. Metabolomics using LC-MS analyses of urine might be a good approach to confirm this hypothesis.
Using an acute mortality test on mice, Saito and Kaneda [14] tried to determine the most toxic fraction isolated from linseed oil heated under drastic conditions. Four components in the isolated fraction were cyclic monomers. The first two reportedly had a disubstituted cyclohexanic structure with two identified ethylenic bonds on a lateral chain, another had a cyclohexyl-1,2-diylidene structure, and the last one presented a disubstituted cyclohexenic structure with an ethylenic bond on one of the chains. However, the most recent studies on the structure of cyclic monomers did not find all the components described by these authors in oil heated under the same conditions.
While reproduction studies have been carried out to evaluate the potential toxicity of heated oils, little work was done on the subject with isolated CFAM fractions. In one experiment [2], female Wistar rats were given a purified diet, including 18% casein and 10% lipids just after mating. The experimental groups contained 1, 0.1, and 0.01% cyclic monomers formed from α-linolenic acid, fed as methyl ester derivatives. A decrease in food intake was observed for the group having 1% CFAM in the diet. The number of pups per litter at birth did not vary among groups. The most spectacular effect observed concerned pup mortality during the first days after birth. In the group containing 1% CFAM, mortality of pups reached 98% during the first three days, and the few survivors did not live longer than five days. This effect was earlier observed by Farmer et al. [15] and Potteau [16] with diets containing equivalent quantities of CFAM present in heated oil.
To verify these results, another reproduction experiment was conducted with a higher amount of protein in the diet (21%) and CFAM were fed as ethyl esters to avoid toxicity of methanol release from methyl esters. High mortality (81%) was found in the group of pups fed heated linseed oil (about 1% CFAM), whereas the same mortality (3%) was found in control group and groups receiving increasing amounts of CFAM (0.2, 0.4, 0.6, and 1% of diet) [17], so this did not confirm the data previously reported. For the group receiving 1% CFAM as ethyl esters, however, the quantity of CFAM found in the livers was about four times lower than what was found in the livers of pups fed the heated oil containing the same quantity of CFAM (1%). These results indicate that ethyl esters of CFAM are not well absorbed and that toxicity of CFAM seems to depend on the form in which these are fed to rats (methyl esters, ethyl esters, triacylglycerols). The same selectivity of incorporation of CFAM (preferential incorporation of 5-membered-ring isomers) was found for animals fed heated oil or purified CFAM fractions, however.
In order to determine if CFAM could have an effect during the gestation period by inducing cellular lesions in the liver of the fetus, Joffre [18] also carried out a reproduction study with increasing amounts of CFAM (0.3, 0.6 and 0.9%) fed as triacylglycerols this time. Feeding female rats with CFAM resulted in a reduction of food intake for those fed the highest quantity of CFAMs (0.9%) as previously described, which resulted in a lower body weight gain while no differences were observed for the average number of fetuses and the mean fetus weight at day 20. The liver weight of fetus from rats fed CFAM was significantly lower than that of the control. Increasing the quantity of CFAM in the diet from 0.3 to 0.9% did not result in any changes in the CFAM content of liver of the fetus, but an accumulation of triacylglycerols was observed in the liver of the fetus in all the groups fed CFAMs. Morphological studies also demonstrated that CFAM formed from linolenic acid induced accumulation of glycogen and changes in mitochondrial ultrastructure. It would then be interesting to determine if these morphological changes would induce any changes in the function of the mitochondria for fatty acid oxidation. If so these changes could explain why at birth mortality could be high when the pup is changing from a diet low in lipids to a diet rich in lipids (maternal milk).
In order to complete the evaluation of the toxicity of CFAM derived from linolenic acid [19], pregnant rats were fed with triacylglycerols containing CFAM (0.7% of diet). These conditions can be considered as physiologically relevant in term of bioavailability of the CFAM, in contrast to those using semipurified fractions of oils, or esters of CFAM. Feeding CFAM significantly reduced the food intake of the females by 20%, without lowering food efficiency. Glycemia of the female rats, the number and weight of the fetus remained unchanged in the CFAM group compared to the control group. In contrast, 1-day-old rats born from females fed with the CFAM diet were significantly lighter than controls (-20%), and their glycemia was reduced by 17%.
A tremendous reduction in insulinemia was observed in the CFAM group compared to the control group, from -47% in females at day 20 of gestation, to -70% in females at parturition and in newborn rats. Furthermore, the results of glycemia and insulinemia in the animals from the restricted group were similar to those of the CFAM group, which suggested that low insulinemia in females and litters in the CFAM group was the consequence of food restriction during gestation and not of endogenous effect of CFAM. Nevertheless, low insulinemia and low weight of the neonates in the CFAM and restricted groups were consistent with the literature [20]. It must be pointed out that hypoinsulinemia and intrauterine growth restriction might predispose the offspring to the development of insulin resistance, cardiovascular disease, obesity, and even overt diabetes in later life [20], which may not be linked to a direct effect of dietary CFAM, however.
In this later work, CFAM, even when fed at high non-nutritional doses and as triacylglycerols, did not induce death of the pups within the first day of life, contradicting previous works done with purified fractions of oils containing CFAM or CFAM-esters, the bioavailability and toxicity per se of which remain questionable. Hypoinsulinemia and low growth of the pups may have been the origin of the adverse effects of the CFAM.
4. Conclusions
CFAM present in heat-treated oils have shown a high intestinal absorption, which depends on their position on the triacylglycerol and on their structure. They may be metabolized, selectively incorporated in tissues and oxidized in a similar way as the precursor essential fatty acids. They also affect the activities of lipid-synthesising and -oxidizing enzymes in rat liver. Based on the data presented in this short review and considering that oxidation and elimination are powerful processes for excretion of CFAM and their low amounts in the diet, we suggest that cyclic fatty acids are not a major concern for human safety when oils are used under normal frying conditions.
References
- Sébédio, J.L. and Juaneda, P. Isomeric and cyclic fatty acids as a result of frying. In: Deep Frying, pp. 57-86 (M.D. Erickson (ed.), AOCS Press, Champaign, IL) (2007).
- Sébédio, J.L. and Grandgirard, A. Cyclic fatty acids: natural sources, formation during heat treatment, synthesis and biological properties. Prog. Lipid Res., 28, 303-336 (1989).
- Combe, N., Constantin, M.J. and Entressangles, B. Absorption intestinale des espèces chimiques nouvelles (E.C.N.) formées lors du chauffage des huiles. Rev. Franç. Corps Gras, 1, 27-28 (1978).
- Martin, J.C., Caselli, C., Broquet, S., Juaneda, P., Nour, M., Sébédio, J.L. and Bernard, A. Effect of cyclic fatty acid monomers on fat absorption and transport depends on their positioning within the ingested triacylglycerols. J. Lipid Res., 38, 1666-1679 (1997).
- Joffre, F., Martin, J., Genty, M., Demaison, L., Loreau, O., Noel, J. and Sébédio, J.L. Kinetic parameters of hepatic oxidation of cyclic fatty acid monomers formed from linoleic and linolenic acids. J. Nutr. Biochem., 12, 554-558 (2001).
- Joffre, F., Roy, A., Bretillon, L., Pasquis, B., Sergiel, J.P., Loreau, O., Chardigny, J.M. and Sébédio, J.L. In vivo oxidation of carboxyl-labelled cyclic fatty acids formed from linoleic and linolenic acids in the rat. Reprod. Nutr. Dev., 44, 123-130 (2004).
- Sébédio, J.L., Chardigny, J.M. and Malpuech-Brugere, C. Physiological effects of trans and cyclic fatty acids. In: Deep Frying, pp. 205-228 (M.D. Erickson (ed.), AOCS Press, Champaign, IL) (2007).
- Ribot, E., Grandgirard, A., Sébédio, J.L., Grynberg, A. and Athias, P. Incorporation of cyclic fatty acid monomers in lipids of rat heart cell cultures. Lipids, 27, 79-81 (1992).
- Crampton, E.W., Common, R.H., Farmer, F.A., Berryhill, F.M. and Wiseblatt, L. Studies to determine the nature of the damage to the nutritive value of some vegetable oils from heat treatment. II. Investigations of the nutritiousness of the products of thermal polymerization of linseed oil. J. Nutr., 44, 177-189 (1951).
- Crampton, E.W., Common, R.H., Farmer, F.A., Wells, A.F. and Crawford, D. Studies to determine the nature of the damage to the nutritive value of some vegetable oils from heat treatment. III. The segregation of toxic and non-toxic material from the esters of heat-polymerized linseed oil by distillation and by urea adduct formation. J. Nutr., 49, 333-346 (1953).
- Iwaoka, W.T. and Perkins, E.G. Metabolism and lipogenic effects of the cyclic monomers of methyl linolenate in the rat. J. Am. Oil Chem. Soc., 55,734-738 (1978).
- Damy Zarambaud, A. and Grandgirard, A. Detoxification by the rat of compounds formed during thermal polymerization of linseed oil. II. Effects of discontinuous administration of the heated oil on the urinary excretion of glucuronides, liver weight and the tissue content of cyclic monomers. Reprod. Nutr. Dev., 2, 409-419 (1981).
- Siess, M.H., Vernevaut, M.F., Grandgirard, A. and Sébédio, J.L. Induction of hepatic drug-metabolizing enzymes by cyclic fatty acid monomers in the rat. Food Chem. Toxicol., 26, 9-13 (1988).
- Saito, M. and Kaneda, T. Studies on the relationship between the nutritive value and the structure of polymerized oils. 11. Mechanisms of toxicity of heat-polymerized oils. Yukagaku, 25, 842-847 (1976).
- Farmer, F.A., Crampton, E.W. and Siddall, M.I. The effect of heated linseed oil on reproduction and lactation in the rat. Science, 113, 408-410 (1951).
- Potteau, B. Influence of heated linseed oil on reproduction in the female rat and on the composition of hepatic lipids in young rats. Ann. Nutr. Aliment., 30, 67-88 (1976).
- Sébédio, J.L., Chardigny, J.M., Juaneda, P., Giraud, M.C., Nour, M., Christie, W.W. and Dobson. G.A. Nutritional impact and selective incorporation of cyclic fatty acid monomers in rats during reproduction. In: 21st World Congress of the International Society for Fat Research. 1995. The Hague, pp. 307-310 (P.J. Barnes, & Associates, Bridgwater).
- Joffre, F. Effets nutritionnels des monomères cycliques issus del'acide α-linolénique chez l'animal. Thesis, Université de Bourgogne: Dijon. p. 153 (2001).
- Bretillon, L., Roy, A., Pasquis, B., and Sébédio, J.L. Dietary cyclic fatty acids derived from linolenic acid do not exhibit toxicity in the rat during gestation. Animal, 2, 1534-1537 (2008).
- Holemans, K., Aerts, L. and Van Assche, F. Fetal growth restriction and consequences for the offspring in animal models. J. Soc. Gynecol. Investig., 10, 392-399 (2003).
In This Section
- Oil Refining
- Action of Natural Antioxidants During Frying
- Formation of New Compounds During Frying - General Observations
- Formation of cyclic fatty acids during frying
- Formation of Epoxy-, Keto- and Hydroxy-Fatty Acids
- Formation of Volatiles and Short-Chain Bound Compounds
- Formation of Dimers and Oligomers
- Oxysterol Formation Frying Oils
- Structural Analysis of the Cyclic Fatty Acids Formed during Frying
- Cyclic Fatty Acids: Isolation and Quantitative Analysis in Food and Biological Tissues
- Analysis of Used Frying Oils and Fats by High-Performance Size-Exclusion Chromatography
- Analysis of Trans Polyunsaturated Fatty Acids
- Determination of Polar Compounds in Used Frying Oils and Fats by Adsorption Chromatography
- Determination of Oxidized Monomeric, Dimeric and Oligomeric Triacylglycerols; Diacylglycerols and Free Fatty Acids
- Separation and Quantification of Oxidized Monomeric, Dimeric and Oligomeric Fatty Acids
- Analysis of Oxidized Fatty Acids
- Analysis of Oxidized Sterols in Frying Oils
- Metabolism and Physiological Effects of Cyclic Fatty Acids Formed from Linoleic and alpha-Linolenic Acids during Frying
- Metabolism and Physiological Effects of Cyclic Fatty Acids Formed from Linoleic and alpha-Linolenic Acids during Frying
- Metabolism of Trans Polyunsaturated Fatty Acids Formed during Frying
- Biological Effects of Frying Oils Mediated by the Activation of Peroxisome Proliferator-Activated Receptors (PPAR)