Plant Triacylglycerol Synthesis
The Authors: Randall Weselake, Jitao Zou and David Taylor
Introduction
Triacylglycerol is composed of three fatty acyl groups esterified to a glycerol backbone at the sn-1, sn-2 and sn-3 positions. In higher plants, triacylglycerol is the predominant component of the oil of the seeds or fruits of oleaginous plants and primarily serves as an energy store to support the growth of the young seedling during the early stages of germination. Triacylglycerol is also an important part of our bio-economy, providing a source of highly reduced carbon for both food and nonfood applications, such as supplying a feedstock for the production of petrochemical alternatives.
The biosynthesis of triacylglycerol occurs in the endoplasmic reticulum (ER) and involves acyl-editing of fatty acyl chains within the nitrogenous phospholipids of the ER. Depending on the plant species, particular reactions of triacylglycerol assembly and acyl-editing may be catalyzed by one or more forms of an enzyme. In addition, in some cases, variants or allelic variants of the same isoenzyme have been identified. Currently, there are no three-dimensional structures available for any of these enzymes. Triacylglycerol droplets accumulate in the outer leaflet of the ER and eventually 'bud off' from the ER to form oil bodies ranging from 0.5 to 2.5 micrometers in diameter which are surrounded by a monolayer of phospholipid, with amphiphilic oil body proteins embedded in the triacylglycerol and phospholipid layer. Although triacylglycerol accumulates to high levels in the seeds and fruits of oleaginous plants, triacylglycerol is also known to collect in other plant parts including pollen and vegetative tissue.
This contribution to the AOCS Lipid Library begins by briefly examining the source of precursors for triacylglycerol assembly. Thereafter, reactions involved in the sequential acylation of the glycerol backbone in the linear part of sn-glycerol-3-phosphate pathway (Kennedy pathway) leading to triacylglycerol are discussed. It is then shown that the fatty acid composition of triacylglycerol can be modified by acyl editing mechanisms in the ER which act on 'diacylglycerol skeletons'. These mechanisms not only affect the diacylglycerol skeleton at the level of phosphatidylcholine (or in some cases, phosphatidylethanolamine), but they also involve acyl-exchange with the acyl-Coenzyme A (CoA) pool. Insights into the details of triacylglycerol biosynthesis, and information on the genes and enzymes involved in this process, have led to innovative strategies to modify the fatty acid composition of triacylglycerol and increase seed oil content. Some of these molecular strategies will be briefly described. Finally, it is shown that leaf tissue can also accumulate small amounts of triacylglycerol but can be encouraged to form more triacylglycerol through various metabolic engineering approaches.
Precursors for Triacylglycerol Biosynthesis
A generalized scheme for triacylglycerol biosynthesis in developing seeds of oleaginous plants is depicted in Figure 1. The de novo biosynthesis of fatty acids occurs in the plastid through the catalytic action of acetyl-CoA carboxylase (EC 6.4.1.2) and the fatty acid synthase complex. Monounsaturated fatty acids from C4 to C18 are formed in the plastid while the fatty acyl chain is attached to the acyl carrier protein of the fatty acid synthase complex. Thioesterase (acyl-acyl carrier protein hydrolase) (EC 3.1.2.14) catalyzes the release of saturated or monounsaturated fatty acyl chain from acyl-acyl carrier protein. In Arabidopsis and many other oilseed species, oleic acid (18:1 Δ9-cis) is a major product of plastidial fatty acid synthesis. Following export from the plastid, fatty acids are re-esterified to CoA to form acyl-CoA through the catalytic action of acyl-CoA synthetase (EC 6.2.1.3) on the outer membrane of the plastid. Further elongation of acyl-CoAs can occur on the ER. For example, in high-erucic-acid rapeseed (Brassica napus), oleoyl-CoA is sequentially elongated to form eicosenoyl (20:1 Δ11-cis)-CoA and erucoyl (22:1 Δ13-cis)-CoA. Both plastidially-exported and -elongated acyl-CoAs constitute the acyl-CoA pool, which can be further modified through acyl-exchange reactions in the ER. Acyl-exchange reactions will be described in more detail later in this contribution.
Figure 1. Generalized scheme for triacylglycerol (TAG) biosynthesis in developing seeds of oleaginous plants. Monounsaturated fatty acids (MUFAs) and saturated fatty acids (SFA) are synthesized in the plastid and, following export from this organelle, are converted to acyl-Coenzyme As (CoAs). Acyl-CoAs can be further elongated through the catalytic action of fatty acid elongase (FAE) on the endoplasmic reticulum (ER). TAG can be synthesized via the acyl-CoA-dependent acylation of the glycerol backbone derived from sn-glycerol-3-phosphate (G3P). Phosphatidic acid phosphatase (PAP) catalyzes the dephosphorylation of phosphatidic acid (PA) to produce sn-1,2-diacylglycerol (DAG) prior to the final acylation catalyzed by acyl-CoA:diacylglycerol acyltransferase (DGAT). DAG can also be converted to phosphatidylcholine (PtdC) via the action of sn-1,2-diacylglycerol:cholinephosphotransferase (CPT) and/or phosphatidylcholine:diacylglycerol cholinephosphotransferase (PDCT). MUFA at the sn-2 position of PtdC can undergo desaturation catalyzed by fatty acid desaturases 1 and 2 (FAD1 and FAD2), respectively. PtdC enriched in polyunsaturated fatty acid (PUFA) can be returned as DAG into the linear part of the G3P pathway leading to TAG via the action of PDCT. Phospholipase A2 (PLA2) may also catalyze the removal of PUFA from the sn-2 position PtdC which in turn is converted to acyl-CoA via the action acyl-CoA synthetase (ACS). In addition, acyl-CoA:lyso-phosphatidylcholine acyltransferase (LPCAT) may catalyze acyl-exchange between the sn-2 position of PUFA-enriched PtdC and the acyl-CoA pool. Low molecular mass soluble acyl-CoA-binding protein (ACBP) may interact with acyl-CoA and encourage the reverse reaction of LPCAT leading to acyl-CoA. Phospholipid:diacylglycerol acyltransferase (PDAT) catalyzes the transfer of fatty acid (FA) at the sn-2 position of PtdC to DAG to also generate TAG. PLA2 and PDAT have been implicated in catalyzing the removal of unusual FAs from the PtdC. LPCAT may catalyze the reacylation of lysophosphatidylcholine (LPC) produced through the action of PLA2 or PDAT. Other abbreviations: GPAT, acyl-CoA:sn-glycerol-3-phosphate acyltransferase; LPA, lysophosphatidic acid; LPAAT, acyl-CoA:lysophosphatidic acid acyltransferase; PC, phosphocholine.
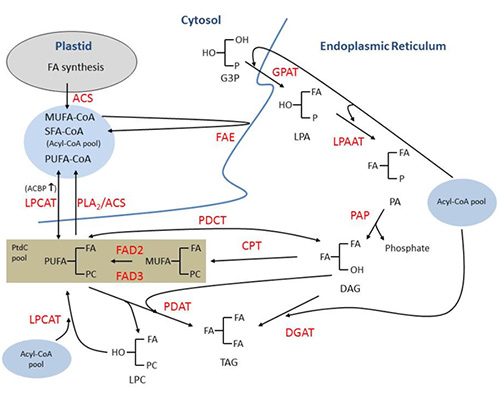
The acyl-CoA pool provides acyl-donor substrate for the acyltransferase reactions of triacylglycerol assembly in the ER. The glycerol backbone for triacylglycerol assembly is in the form of sn-glycerol-3-phosphate, which is generated through the catalytic action of sn-glycerol-3-phosphate dehydrogenase (EC 1.1.1.8). The dihydroxyacetone phosphate substrate for this reaction is derived from glycolysis.
Triacylglycerol Can Be Synthesized through Sequential Acylation of the Glycerol Backbone
Triacylglycerol can be formed through three sequential acyl-CoA-dependent acylations of the glycerol backbone beginning with sn-glycerol-3-phosphate. Removal of the phosphate group occurs prior to the final acylation. The acylation of sn-glycerol-3-phosphate is catalyzed by acyl-CoA:sn-glycerol-3-phosphate acyltransferase (GPAT; EC 2.3.1.15). The second acylation is catalyzed by acyl-CoA:lyso-phosphatidic acid acyltransferase (LPAAT; EC 2.3.1.51). Phosphatidic acid phosphatase (PAP EC 3.1.3.4) catalyzes the removal of the phosphate group from phosphatidic acid to generate sn-1,2-diacylglycerol. The final acyl-CoA-dependent acylation is catalyzed by acyl-CoA:diacylglycerol acyltransferase (DGAT; EC 3.2.1.20).
Acylation of sn-glycerol-3-phosphate mediated by GPAT to form lyso-phosphatidic acid initiates the synthesis of all glycerolipid molecules. The acyl-CoA substrate preferences of GPAT contribute to the asymmetric distribution of saturated and unsaturated fatty acids in the sn-1 and sn-2 positions of phospholipid molecules as well as triacylglycerols. Considered as a potential 'bottleneck' step in the flow of carbon into triacylglycerol since originally defined by Eugene Kennedy, it is now realized that GPAT-mediated regulation of glycerolipid synthesis is more complex than previously thought. Genes encoding for ER-bound GPAT were first cloned from Saccharomyces cerevisiae. Interestingly, over- expressing one isoform (SCT1) in yeast leads to enhanced accumulation of triacylglycerol, whereas increased expression of another (GPT1) results in decreased level of storage lipids. Higher plants also possess homologs of the yeast GPATs, and members of the Arabidopsis GPAT gene family displayed GPAT activity when expressed in yeast. But mutations in Arabidopsis GPAT genes do not appear to impact seed oil level, and recent findings have indicated that Arabidopsis GPAT4 and -6 have GPAT activity with sn-2 preference and participate in cutin biosynthesis through generating monoacylglycerol.
The second acylation step catalyzed by LPAAT appears to be a reaction which is specialized in certain oilseeds. For example, in oilseed rape (B. napus), sunflower (Helianthus annuus) and flax (Linum usitatissimum), the acyl-CoA selectivity of LPAAT favours oleoyl-, linoleoyl (18:2 Δ9,12-cis)- and α-linolenoyl (18:3 Δ9,12,15-cis)-CoAs, respectively. In high-erucic-acid rapeseed and other Brassicaceae, the LPAAT discriminates against very long chain fatty acyl-CoAs such as eicosenoyl-, erucoyl- and nervonoyl (24:1 Δ15-cis)-CoAs. LPAATs from Tropaeolaceae and Limnanthaceae, in contrast, show a strong affinity for very-long-chain fatty acyl-CoAs. In other oil crops, such as coconut and palm, the resident LPAATs favour the incorporation of medium-chain fatty acids from decanoic (10:0) to myristic (14:0) acid.
PAP1, which catalyzes the dephosphorylation of phosphatidic acid, is a Mg2+-dependent enzyme associated with triacylglycerol biosynthesis. In contrast, PAP2 does not require Mg2+ and can utilize a range of phosphate-containing substrates including phosphatidic acid. Although PAP1 exists in soluble form, oleate has been shown to cause PAP1 activity to associate with membranes in developing safflower cotyledons. Recent studies with Arabidopsis have demonstrated that PAP1 and PAP2 are involved in the eventual provision of diacylglycerol from the the eukaryotic pathway for galactolipid synthesis in the plastid, similar to yeast
Two families of membrane-bound DGAT have been identified, both utilizing long-chain acyl-CoAs but sharing essentially no homology. Hydropathy plot analyses suggest that DGAT1 has several trans-membrane segments whereas DGAT2 may only have a few. DGAT2, however, appears to have a role in catalyzing the incorporation of unusual fatty acids such as ricinoleic acid (12-OH 18:1 Δ9-cis), lesquerolic acid (14-OH 20:1 Δ11-cis), eleostearic acid (18:3 Δ9-cis,11-trans,13-trans) and epoxy fatty acids into triacylglycerol. Localization studies using confocal microscopy suggest that DGAT1 and DGAT2 are localized in different regions of the ER. A soluble form of DGAT has also been identified in developing peanut (Arachis hypogaea) cotyledons. More recently, a membrane-bound DGAT with preference for acetyl-CoA was identified in burning bush (Eunoymus alatus). The sn-3 acetyl-triacylglycerol produced via this enzyme exhibits decreased viscosity and may be useful in certain biodiesel applications directly, without conversion to methyl esters.
Acyl Editing in the Endoplasmic Reticulum Can Affect the Fatty Acid Composition of Triacylglycerol
Polyunsaturated fatty acids are mainly formed using phosphatidylcholine as a substrate for the catalytic action of membrane-bound fatty acid desaturase 2 and 3 (FAD2 and FAD3), which leads to the conversion of oleoyl moieties to linoleoyl and linolenoyl, respectively (see the web page on fatty acid biosynthesis). The formation of unusual fatty acids, such as ricinoleic and epoxy fatty acids, also occurs on phosphatidylcholine and involves the action of enzymes which are evolutionarily related to desaturases. In some plant species, however, polyunsaturated fatty acid formation can also occur through the action of desaturases that use acyl-CoA as substrates. Polyunsaturated fatty acids formed on phosphatidylcholine are eventually incorporated into triacylglycerol via a number of possible routes.
Kinetically distinct pools of sn-1, 2-diacylglycerol have been shown to exist within the ER of developing soybean cotyledons. sn-1,2-Diacylglycerol produced through the action of PAP was used in the production of phosphatidylcholine whereas sn-1,2-diacylglycerol used by DGAT for triacylglycerol assembly was derived from phosphatidylcholine. Diacylglycerol:cholinephosphotransferase (CPT; EC 2.7.8.2) can catalyze the formation of phosphatidylcholine from sn-1,2-diacylglycerol and CDP-choline. Although the reverse reaction of CPT has been implicated in moving the sn-1,2-diacylglycerol skeleton from phosphatidylcholine back into sn-1,2-diacylglycerol, the reaction kinetics are highly unfavourable in this direction. More recently, another enzyme with an apparent role in sn-1,2-diacylglycerol/phosphatidylcholine interconversion was identified in Arabidopsis. Phosphatidylcholine:diacylglycerol cholinephosphotransferase (PDCT) catalyzes the transfer of phosphocholine from phosphatidylcholine to sn-1,2-diacylglycerol and can serve as a reversible route for moving sn-1,2-diacylglycerol-containing oleoyl moieties into phosphatidylcholine and removing diacylglycerol-containing polyunsaturated fatty acids from phosphatidylcholine. In turn, sn-1,2-diacylglycerol enriched in polyunsaturated fatty acids can be used by DGAT in triacylglycerol assembly.
Phospholipid:diacylglycerol acyltransferase (PDAT; EC 2.3.1.158) catalyzes the acyl-CoA-independent synthesis of triacylglycerol using phosphatidylcholine as an acyl donor and sn-1,2-diacylglycerol as an acyl acceptor. This reaction may be useful in transferring unusual fatty acids from phosphatidylcholine into triacylglycerol. The membrane anchor from yeast PDAT can be removed to produce a soluble enzyme displaying PDAT activity. This solubilized enzyme can also, to some extent, catalyze the transfer of acyl moieties between two molecules of sn-1,2-diacylglycerol to generate triacylglycerol and monoacylglycerol. PDAT1 and DGAT2 have overlapping functions in triacylglycerol biosynthesis in developing seeds and pollen of Arabidopsis. However, the degree to which each enzyme contributes to triacylglycerol biosynthesis in developing seeds of Arabidopsis or in other oilseeds is still undefined.
Phospholipase A2 (PLA2; EC 3.1.1.4) has also been implicated as a possible way of removing unusual fatty acyl moieties from phosphatidylcholine. In this latter case, acyl-CoA synthetase would be required to convert the released free fatty acid into acyl-CoA, which could in turn serve as a substrate for the acyl-CoA-dependent acyltransferases.
Acyl-CoA:lysophosphatidylcholine acyltransferase (LPCAT; EC 2.3.1.23) represents yet another possible route for moving polyunsaturated fatty acids or unusual fatty acids from phosphatidylcholine into the acyl-CoA pool. The forward reaction catalyzed by LPCAT uses acyl-CoA and lyso-phosphatidylcholine as co-substrates to form phosphatidylcholine and free CoA. The reverse reaction leading to lyso-phosphatidylcholine and acyl-CoA, however, is highly unfavourable. Acyl-cycling between phosphatidylcholine and lyso-phosphatidylcholine may be primarily controlled by a coordinated forward acylation to produce phosphatidylcholine catalyzed by LPCAT and the reverse deacylation of phosphatidylcholine and resynthesis of acyl-CoA via PLA2 and acyl-CoA synthetase, respectively. Additionally there is strong evidence that acyl-CoA binding proteins, which interact with acyl-CoA, may serve to reduce the free concentration of acyl-CoA and thus encourage the reverse reaction of LPCAT to occur. Results of in vitro assays using microsomal fractions from yeast containing recombinant Arabidopsis LPCAT and soluble low molecular mass acyl-CoA-binding protein (from B. napus) suggest that the combined forward and reverse reactions of LPCAT facilitate acyl exchange between the sn-2 position of phosphatidylcholine and the acyl-CoA pool.
Metabolic Engineering of Triacylglycerol Biosynthesis
Modification of the fatty acid composition of major oilseed crops has been achieved through manipulation of enzyme-catalyzed reactions in lipid biosynthesis (see also our web page on production of unusual fatty acids). In many instances, reactions in both fatty acid and triacylglycerol biosynthesis were modified via the introduction of cDNAs encoding enzymes with desired substrate selectivities/specificities from other sources. A major obstacle to producing industrially useful fatty acids in Brassica oilseed species remains the inability of the resident LPAAT to incorporate these fatty acyl groups at the sn-2 position of the glycerol backbone. Triacylglycerols enriched in lauric acid (12:0) can serve as feedstock for the detergent industry. Researchers at Calgene introduced a lauroyl-ACP thioesterase from the California bay laurel (Umbellularia californica) into B. napus to facilitate production of lauric acid and its subsequent incorporation into seed triacylglycerol. Incorporation of lauroyl moieties, however, was limited to the sn-1 and sn-3 positions of the triacylglycerol of the transgenics because the endogenous B. napus LPAAT discriminated against saturated acyl-CoA substrates. This obstacle was overcome by co-expressing, in B. napus, the cDNA encoding the California bay lauroyl-ACP thioesterase in combination with a cDNA encoding an LPAAT from coconut (Cocus nucifera). The endogenous LPAAT of high-erucic-acid B. napus also discriminates against erucoyl moieties. In order to overcome this problem, biotechnologists have transformed high-erucic-acid B. napus with cDNAs encoding LPAATs from other species which are capable of incorporating erucoyl moieties at the sn-2 position.
Although the introduction of castor bean (Ricinus communis) fatty acid hydroxylase led to the production of some ricinoleic acid in Arabidopsis, the content of this unusual fatty acid was further increased in the seed triacylglycerol through co-expression of cDNA encoding DGAT2 from castor bean which exhibited a preference for substrates containing ricinoleic acid. Ultra-high levels of incorporation of unusual fatty acids, however, have not been achieved in genetic engineering of major oilseed crops. In contrast, castor bean contains seed triacylglycerol with greater than 90% ricinoleic acid. Garden nasturtium (Tropaeolum majus), despite having only 15% oil, has triacylglycerol with 75% erucic acid. Somehow these species are capable of orchestrating lipid synthesis to produce oil that is highly enriched in one type of fatty acid making the oil more appealing as an industrial feedstock. Work continues to try to mimic the production of such highly homogeneous triacylglycerols in major oilseed crops through biotechnology.
Reactions in triacylglycerol biosynthesis have also been manipulated to increase seed oil content. A number of studies have suggested that, in certain plant species, the level of DGAT activity during seed development may have a substantial effect on the flow of carbon into seed oil. During development of the zygotic embryo in B. napus, sn-1,2-diacylglycerol was shown to be the next most prevalent metabolite in the sn-glycerol-3-phosphate pathway following triacylglycerol, suggesting that the DGAT-catalyzed reaction represented a 'bottleneck' in the flow of carbon into seed oil. In addition, the activity of DGAT in microsomes from developing embryos of B. napus is considerably less than that of GPAT, LPAAT or PAP. Overexpression of cDNAs encoding either Arabidopsis DGAT1 or a variant of B. napus DGAT1 during seed development in B. napus resulted in increased seed oil content under both greenhouse and field conditions. This was the first example of how DGAT overexpression was used to increase seed oil content in a major oilseed crop.
Top-down control analysis of carbon flux has been used to investigate the relative contributions of fatty acid biosynthesis versus triacylglycerol biosynthesis to oil accumulation in various plant species. The reactions of fatty acid biosynthesis make up Block A whereas the reactions of triacylglycerol assembly make up Block B, with acyl-CoA linking the two Blocks. In developing zygotic embryos of B. napus L. cv Westar, about 70% of the control in carbon flux leading to triacylglycerol was associated with Block B suggesting that metabolic engineering of triacylglycerol assembly to reduce the level of control in Block B could potentially boost the triacylglycerol content of the zygotic embryo. Transgenic B. napus L. cv Westar overexpressing a cDNA encoding B. napus DGAT1 exhibited only about 50% control in Block B (a 20% reduction) which was probably associated with increased DGAT activity during seed development. Thus, metabolic flux analysis can be used as a tool for informed metabolic engineering. In another study, a specific allele of DGAT1 in maize (Zea mays) has been associated with both increased embryo oil content and oleic acid content in the oil. A cDNA encoding a DGAT2 from a fungal source was also used to increase the oil content of soybean (Glycine max) cotyledons. A high-throughput procedure for screening for DGAT variants with increased activity has recently been developed, which may eventually provide improved DGATs for further increasing seed oil content.
Other metabolic engineering strategies targeted at increasing the triacylglycerol content of seeds have involved manipulation of other reactions in carbon flow that provide building blocks for fatty acid and storage lipid biosynthesis. There has also been considerable interest in modifying the action of transcription factors that can up-regulate a series of biochemical reactions in carbon flow. Transcription factors can interact with the promoter regions of genes encoding enzymes catalyzing reactions in carbon flow. Thus far, the transcription factors identified appear to regulate the expression of enzymes involved in fatty acid biosynthesis and other aspects of carbon flow rather than the expression of enzymes involved in triacylglycerol assembly. Modification of the expression of genes encoding transcription factors through genetic engineering, however, is often accompanied by pleiotropiceffects that can negatively affect plant development.
Triacylglycerol Biosynthesis in Vegetative and Root Tissue
Numerous plant species have been shown to contain relatively low levels of triacylglycerol in the cytosol of leaves. Labelling experiments with 14C-acetate have suggested that leaf triacylglycerol may serve as a diurnal photosynthetic store, similar to starch. Seven- to 20-fold increases in the triacylglycerol content of leaves (Nicotiana tabacum and Arabidopsis) have been achieved through overexpression of DGAT1. Leaf triacylglycerol content has also been increased by blocking β-oxidation of leaf fatty acids and through ectopic expression of LEAFYCOTYLEDON 2 which encodes a transcription factor associated with embryo maturation. Another innovative approach to producing triacylglycerol droplets in Arabidopsis leaves was based on the inactivation of a human homolog of CGI-58. In humans, this protein aids lipolysis on lipid droplets. A defect in the encoding gene leads to the abnormal intracellular deposition of lipid droplets in a number of tissue types in a disorder known as Chanarin-Dorfman Syndrome.
Recently, a soluble DGAT was identified in Arabidopsis leaves that appears to have a role in surface cutin formation. Presumably, the enzyme participates in the formation of triacylglycerol droplets, which may serve as a reserve of fatty acyl moieties for cutin biosynthesis. The possibility of producing high levels of triacylglycerol in the leaves of high biomass-producing plants under field conditions may eventually contribute to a source of triacylglycerols for biofuel and other petrochemical alternatives, which do not compromise the availability of seed oils for food and feed.
Tetraena mongolica Maxim is an example of a plant species that contains substantial triacylglycerol in its stem tissue. The plant is often called ‘oil firewood’. The phloem cells contain cytoplasmic oil bodies which contain about 9% (dry weight) triacylglycerol. The stem appears to have evolved as an organ for storing carbon in support of the development of new shoots later in the year.
There are also examples of triacylglycerol accumulation in roots. Yellow nutsedge (Manihot esculenta) accumulates approximately 25% oil and 30% starch in the tubers. Thus, the plant may be an attractive system in which to study mechanisms associated with the partitioning of carbon between oil and starch. In addition, the pkl mutant of Arabidopsis produces triacylglycerol in its roots with a fatty acid composition similar to its seed oil.
References
- Andrianov, V., Borisjuk, N., Pogrebnyak, N., Brinker, A., Dixon, J., Spitsin, S., Flynn, J., Matyszczuk, P., Andryszak, K., Laurelli, M., Golovkin, M. and Koprowski, H. Tobacco as a production platform for biofuel: overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation and shifts the composition of lipids in green biomass. Plant Biotechnol. J., 8, 277-287 (2010) (DOI: 10.1111/j.1467-7652.2009.00458.x).
- Bates, P.D., Durrett, T.P., Ohlrogge, J.B. and Pollard, M. Analysis of acyl fluxes through multiple pathways of triacylglycerol synthesis in developing soybean embryos. Plant Physiol., 150, 55-72 (2009) (DOI: 10.1104/pp.109.137737).
- Baud, S. and Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res., 49, 235-249 (2010) (DOI: 10.1016/j.plipres.2010.01.001).
- Burgal, J., Shockey, J., Lu, C.F., Dyer, J., Larson, T., Graham, I. and Browse, J. Metabolic engineering of hydroxyl fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol. J., 6, 819-831 (DOI: 10.1111/j.1467-7652.2008.00361.x).
- Carlsson, A.S. Plant oils as feedstock alternatives to petroleum – A short survey of potential oil crop platforms. Biochimie, 91, 665-670 (2009) (DOI: 10.1016/j.biochi.2009.03.021).
- Dahlqvist, A., Ståhl, U., Lenman, M., Banas, A., Lee, M., Sandager, L. Ronne, H. and Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Nat. Acad. Sci. U.S.A., 97, 6487-6492 (2000).
- Durrett, T.P., Benning, C. and Ohlrogge, J. Plant triacylglycerols as feedstocks for the production of biofuels. Plant J., 54, 593-607 (2008) (DOI: 10.1111/j.1365-313X.2008.03442.x).
- Durrett, T.P., McClosky, D.D., Tumaney, A.W., Elzinga, D.A., Ohlrogge, J. and Pollard, M. A distinct DGAT with sn-2 acetyltransferase activity that synthesizes unusual, reduced-viscosity oils in Euonymus and transgenic seeds. Proc. Nat. Acad. Sci. U.S.A., 107, 9464-9469 (2010) (DOI: 10.1073/pnas.1001707107).
- Eastmond, P.J., Quettier, A.-L., Kroon, J.T.M., Craddock, C., Adams, N. and Slabas, A.R. PHOSPHATIDIC ACID PHOSPHOHYDROLASE1 and 2 regulate phospholipid synthesis at the endoplasmic reticulum in Arabidopsis. Plant Cell, 22, 2796-2811 (2010) (DOI: 10.1105/tpc.109.071423).
- Ghosal, A., Banas, A., Ståhl, U., Dahlqvist, Lindqvist, Y. and Stymne, S. Saccharomyces cerevisiae phospholipid:diacylglycerol acyltransferase (PDAT) devoid of its membrane anchor region is a soluble and active enzyme retaining its substrate specificities. Biochim. Biophys. Acta – Mol. Cell Biol. Lipids, 1771, 1457-1463 (2007) (DOI: 10.1016/j.bbalip.2007.10.007).
- Harwood, J.L. Fatty acid biosynthesis. In: Plant Lipids: Biology, Utilization and Manipulation, pp. 27-66 (D.J. Murphy (ed), Blackwell Publishing, Oxford) (2005).
- Lardizabal, K., Effertz, R., Levering, C., Mai, J., Pedroso, M.C., Jury, T., Aasen, E., Gruys, K. and Bennett, K. Expression of Umbelopsis ramanniana DGAT2A in seed increases oil in soybean. Plant Physiol. 148, 89-96 (2008) (DOI: 10.1104/pp.108.123042).
- Li-Beisson, Y., Shorrosh, B., Beisson, F., Andersson, M.X., Arondel, V., Bates, PD., Baud, S., Bird, D., DeBono, A., Durrett, T.P., Franke, R.B., Graham, I.A., Katayama, K., Kelly, A.A., Larson, T., Markham, J.E., Miquel, M., Molina, I., Nishida, I., Rowland, O., Samuels, L., Schmid, K.M., Wada, H., Welti, R., Xu, C., Zallot, R. and Ohlrogge, J. Acyl-lipid metabolism. In: The Arabidopsis Book, published online (http://www.bioone.org/doi/pdf/10.1199/tab.0133).
- Lin, W. and Oliver, D.J. Role of triacylglycerols in leaves. Plant Sci., 75, 233-237 (2008) (DOI: 10.1016/j.plantsci.2008.04.003).
- Lu, C., Xin, Z., Miquel, M. and Browse, J. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proc. Nat. Acad. Sci. U.S.A., 106, 18837-18842 (2009) (DOI: 10.1073/pnas.0908848106).
- Lung, S.C. and Weselake, R.J. Diacylglycerol acyltransferase: A key mediator of plant triacylglycerol synthesis. Lipids, 41,1073-1088 (2006).
- Maisonneuve, S., Bessoule, J.J., Lessire, R., Delseny, M. and Roscoe, T.J. Expression of rapeseed microsomal lysophosphatidic acid acid acyltransferase isozymes enhances seed oil content in Arabidopsis. Plant Physiol., 152, 670-684 (2010) (DOI: 10.1104/pp.109.148247).
- Nakamura, Y., Koizumi, R., Shui, G., Shimojima, M., Wenk, M.R., Ito, T. and Ohta, H. Arabidopsis lipins mediate eukaryotic pathway of lipid metabolism and cope critically with phosphate starvation. Proc. Nat. Acad. Sci. U.S.A. 106, 20978-20983 (2009) (DOI: 10.1073/pnas.0907173106).
- Rani, S.H., Krishna, T.H.A., Saha, S., Negi, A.S. and Rajasekharan, R. Defective in cuticular ridges (DCR) of Arabidopsis thaliana, a gene associated with surface cutin formation, encodes a soluble diacylglycerol acyltransferase. J. Biol. Chem., 285, 38337-38347 (2010) (DOI: 10.1074/jbc.M110.133116).
- Saha, S., Enugutti, B., Rajakumari, S. and Rajasekharan, R. Cytosolic triacylglycerol biosynthetic pathway in oilseeds. Molecular cloning and expression of peanut cytosolic diacylglycerol acyltransferase. Plant Physiol. 141, 1533-1543 (2006) (DOI: 10.1104/pp.106.082198).
- Siloto, R.M.P., Truksa, M., He, X.H., McKeon, T. and Weselake, R.J. Simple methods to detect triacylglycerol biosynthesis in a yeast-based recombinant system. Lipids, 44, 963-973 (2009) (DOI: 10.1007/s11745-009-3336-0).
- Slocombe, S.P., Cornah, J., Pinfield-Wells, H., Soady, K., Zhang, Q.Y., Gilday, A., Dyer, J.M. and Graham, I.A. Oil accumulation in leaves directed by modification of fatty acid breakdown and lipid synthesis pathways. Plant Biotechnol. J., 7, 694-703 (2009) (DOI: 10.1111/j.1467-7652.2009.00435.x).
- Snyder, C.L., Yurchenko, O.P., Siloto, R.M.P., Chen, X., Liu, Q., Mietkiewska, E. and Weselake, R.J. Acyltransferase action in the modification of seed oil biosynthesis. New Biotechnol., 26, 11-16 (2009) (DOI: 10.1016/j.nbt.2009.05.005).
- Wang, G., Lin, Q. and Xu, Y. Tetraena mongolica Maxim can accumulate large amounts of triacylglycerol in phloem cells and xylem parenchyma stems. Phytochemistry, 68, 2112-2117 (2007) (DOI: 10.1016/j.phytochem.2007.04.040).
- Weselake, R.J. Storage lipids. In: Plant Lipids: Biology, Utilization and Manipulation, pp. 162-221 (D.J. Murphy (ed), Blackwell Publishing, Oxford) (2005).
- Weselake, R.J., Shah, S., Tang, M., Quant, P.A., Snyder, C.L., Furukawa-Stoffer, T.L., Zhu, W., Taylor, D.C., Zou, J., Kumar, A., Hall, L., Laroche, A., Rakow, G., Raney, P., Moloney, M.M. and Harwood, J.L. Metabolic control analysis is helpful for informed genetic manipulation of oilseed rape (Brassica napus) to increase seed oil content. J. Exp. Bot., 59, 3543-3549 (2008) (DOI: 10.1093/jxb/ern206).
- Weselake, R.J., Taylor, D.C., Rahman, M.H., Shah, S., Laroche, A., McVetty, P.B.E. and Harwood, J.L. Increasing the flow of carbon into seed oil. Biotechnol. Adv., 27, 866-878 (2009) (DOI: 10.1016/j.biotechadv.2009.07.001).
- Xu, J., Francis, T., Mietkiewska, E., Giblin, E.M., Barton, D.L., Zhang, Y., Zhang, M. and Taylor, D.C. Cloning and characterization of an acyl-CoA-dependent diacylglycerol acyltransferase 1(DGAT1) gene from Tropaeolum majus, and a study of the functional motifs of the DGAT protein using site-directed mutagenesis to modify enzyme activity and oil content. Plant Biotechnol. J., 6, 799-818 (2008) (DOI: 10.1111/j.1467-7652.2008.00358.x).
- Yang, W,, Pollard, M,, Li-Beisson, Y,, Beisson, F., Feig, M. and Ohlrogge, J. A distinct type of glycerol-3-phosphate acyltransferase with sn-2 preference and phosphatase activity producing 2-monoacylglycerol. Proc. Nat. Acad. Sci. U.S.A., 107, 12040-12045 (2010) (DOI: 10.1073/pnas.0914149107).
- Yurchenko, O.P. and Weselake, R.J. Involvement of low molecular mass soluble acyl-CoA-binding protein in seed oil biosynthesis. New Biotechnol., 28, 97-109 (2011) (DOI: 10.1016/j.nbt.2010.09.011)
- Zhang, M., Fan., J., Taylor, D.C. and Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell, 21, 3885-3901 (2009) (DOI: 10.1105/tpc.109.071795).
- Zheng, Z. and Zou, J. The initial step of the glycerolipid pathway: identification of glycerol 3-phosphate/dihydroxyacetone phosphate dual substrate acyltransferases in Saccharomyces cerevisiae. J. Biol. Chem., 276, 41710-41716 (2001) (DOI: 10.1074/jbc.M104749200).
- Zheng, Z., Xia, Q., Dauk, M., Shen, W., Selvaraj, G., and Zou, J. Arabidopsis AtGPAT1, a member of the membrane-bound glycerol-3-phosphate acyltransferase gene family, is essential for tapetum differentiation and male fertility. Plant Cell, 15, 1872-1887 (2003) (DOI: 10.1105/tpc.012427).
- Zheng, P., Allen, W.B., Roesler, K., Williams, M.E., Zhang, S., Li, J., Glassman, K., Ranch, J., Nubel, D., Solawetz, W., Bhattramakki, D., Llaca, V., Deschamps, S., Zhong, G.-Y., Tarczynski, M.C. and Shen, B. A phenylalanine in DGAT is a key determinant of oil content and composition in maize. Nat. Gen., 40, 367-372 (2008) (DOI: 10.1038/ng.85).
In This Section
- Plant Fatty Acid Synthesis
- Production of Unusual Fatty Acids in Plants
- Arabidopsis Acyl-Coenzyme A-Binding Proteins
- Long Chain acyl-coA Synthetases and Other Acyl Activating Enzymes
- Plant Triacylglycerol Synthesis
- Triacylglycerol Biosynthesis in Eukaryotic Microalgae
- Subcellular Oil Droplets and Oleosins in Plants
- Triacylglycerol Mobilisation in Plants
- Role of Transcription Factors in Storage Lipid Accumulation in Plants
- Biosynthesis of Plant Lipid Polyesters
- Rubber Biosynthesis in Plants
- Carotenoid Biosynthesis and Regulation in Plants
- The Oxylipin Biosynthetic Pathways in Plants
- N-Acylphosphatidylethanolamines (NAPEs), N-acylethanolamines (NAEs) and Other Acylamides: Metabolism, Occurrence and Functions in Plants
- Phosphoinositide Signaling in Plants
- Plant Lipidomics
- 50 years of Galactolipid Research: The Beginnings
- Transport and function of lipids in the plant phloem