Subcellular Oil Droplets and Oleosins in Plants
The Author: Anthony H.C. Huang
1. Introduction
Eukaryotes and prokaryotes contain neutral lipids in subcellular droplets for food reserves and other purposes. These lipid droplets are present in seeds, flowers, pollen and fruits of higher plants; the vegetative and reproductive organs of primitive plants, algae, fungi, and nematodes; mammalian organs/tissues such as mammalian glands and adipose tissues; and bacteria. Of all these lipid droplets, the oil bodies (OBs) in seeds are the most prominent and were studied extensively early on [1-6].
Seeds of most plant species store oils (triacylglycerols (TAGs)) as a food reserve for germination and post-germinative growth (there is a separate web page on triacylglycerol biosynthesis). The TAGs are present in small subcellular spherical OBs of approximately 0.5 to 1 μm in diameter. Each OB has a matrix of TAGs surrounded by a layer of phospholipids (PLs) and structural proteins termed oleosins. The small size of OBs provides a large surface area per unit TAG, which would facilitate lipase binding and lipolysis during germination. OBs inside the cells of mature seeds or in isolated preparations are highly stable and do not aggregate or coalesce. This stability is in contrast to the instability of artificial liposomes made from amphipathic and neutral lipids; liposomes generated via sonication will gradually coalesce. Also, lipid droplets in yeast and special mammalian cells, as well as extracellular lipoproteins in mammals and insects, are unstable because they undergo dynamic metabolic fluxes of their surface and matrix constituents [5,7,8]. Seed OBs are stable because their surface is shielded by a layer of oleosins, which, for firm anchorage, has a long hydrophobic stretch that is absent in proteins on lipid droplets of other organisms. In maturing seeds, TAGs, PLs and oleosins are synthesized in endoplasmic reticulum (ER), from which budding OBs are released.
2. Oleosins, the Structural Proteins on Oil Bodies
Oleosins in seeds are small proteins of about 15-26 kDa [1,6]. They completely cover the surface of the subcellular OB (Fig. 1). They can be abundant in seeds with a high proportion of oils and small OBs (therefore large total OB surface area). For example, Arabidopsis seed have more than 40% (wt/wt) oils and small OBs of ~0.5 μm diameter, and 10% of the seed proteins are oleosins.
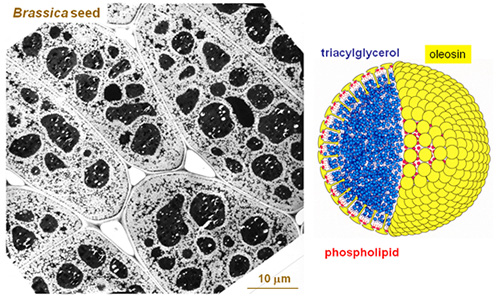
Figure 1. Oil bodies in seed. Left panel is a transmission electron micrograph of a Brassica seed, showing large and conspicuous storage protein bodies and small but numerous oil bodies. Right panel shows a model of an oil body, which includes a matrix of triacylglycerols (in blue) enclosed by a layer of phospholipids (red) and the structural protein oleosin (yellow). The three types of molecules are drawn to similar scales, whereas the diameter of the OB has been reduced 24 times to magnify the surface structure.
Hundreds of genes encoding oleosins have been identified in diverse plant species, ranging from higher to primitive plants including the moss Physcomitrella [9]. Non-plant organisms do not have oleosins, and oleosin-like proteins may be present in algae. Arabidopsis has 17 genes encoding oleosins: 5 active in seed, 3 active in both seed and pollen, and 9 (8 in tandem) active in the floral tapetum cells [10]. Minor proteins present in isolated seed OB fractions of some species have been termed caleosin and steroleosin [4]. They do not have a long hydrophobic sequence, even though they have a short sequence similar to but much less conserved than the proline knot sequence in oleosins (to be described). Their mode of association with OBs should not be similar to that of oleosins, and their function and the possibility of their being remnants of ER after biogenesis need to be explored.
An oleosin molecule can be divided into 3 portions according to its amino acid sequence (Fig. 2). The N-terminal portion can be short or long (e.g., 6-68 residues in Arabidopsis) and hydrophilic or amphipathic. The central portion is a long uninterrupted hydrophobic stretch of 72 residues. The C-terminal portion can be short or very long (e.g. 28-1,000 residues in Arabidopsis oleosins), and its ~40 residues adjacent to the central hydrophobic stretch can form an amphipathic α-helical structure that interacts horizontally with the charged phosphate and choline groups of the PL layer on the OB surface. The very long polypeptide of the C-terminal portion beyond these 40 residues may have little functional significance.
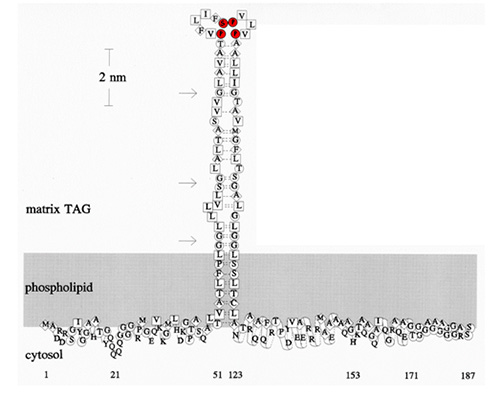
Figure 2. A model of maize 18-kDa oleosin on the surface of an oil body. Shaded area represents the PL layer with the head groups facing the cytosol. Symbols of amino acid residues and their hydropathy indexes: square for very hydrophobic, diamond for hydrophobic, circle for amphipathic, and no enclosure for hydrophilic residues. The N-terminus (residues 1-50) is amphipathic. The central hydrophobic hairpin (residues 51-122) is depicted as anti-parallel β-strands with a proline knot at the loop, which includes 3 proline and 1 serine residues (shown in red). The anti-parallel strands are aligned according to the best-fit pairing of amino acids of similar hydropathic indexes; two dotted lines represent an identical fit, and one dotted line denotes a close fit. The strands may fold and curl at the more flexible glycine or serine pairs (indicated by arrows) to form more bonding. Although the hairpin arms are depicted to be anti-parallel β-strands, they could form anti-parallel α-helices instead. The major portion of the C-terminus (residues 123-170) is an amphipathic α-helical structure interacting with the PL surface horizontally. The C-terminal extension residues (171-187) may have no specific function; the length of this C-terminal extension and its residues are highly variable in oleosins of diverse plant species.
The central hydrophobic stretch of 72 uninterrupted nonpolar residues is the hallmark of an oleosin. No other protein in any organism has such a long, or even half as long, hydrophobic stretch. Proteins on the surface of extracellular or intracellular lipid droplets, such as apolipoproteins, perilipin, adipophilin, and caveolin in mammals, phasin in bacteria and lipid-associated protein (PAP) in plastids, do not have a long hydrophobic stretch [7,8]; their polypeptides run parallel to the surface of rather than penetrate into the lipid droplets. The 72-residue hydrophobic stretch of an oleosin is sufficiently long (a trans-membrane [PL bilayer] peptide has ~20-25 residues) to form a hairpin that penetrates the surface PL monolayer of an OB into the matrix (Figs. 1 and 2). The center of the hydrophobic stretch has 3 proline residues and 1 serine residue that could interact among themselves to form a “proline knot” in a highly hydrophobic environment (Fig. 2). The formation of the proline knot could allow for creation of a hairpin structure of the whole hydrophobic stretch, with two arms of 30 residues each joined by a turn of 12 residues. The 72 residues of the hydrophobic stretch are conserved in terms of hydrophobicity among oleosins of diverse species, and the conservation is higher at the proline knot and its immediate vicinity (-PX5SPX3P-, X being a nonpolar residue). All oleosins contain the 3 proline and 1 serine residues at identical locations in the center of the hydrophobic stretch.
Although the hairpin structure of the central hydrophobic portion of an oleosin molecule has been well established, the secondary structures (α or β) of the loop and the 2 arms remain uncertain [6]. The uncertainty lies in difficulties in creating an environment of a matrix of TAGs covered with a monolayer of PLs to which oleosins can be added and then in measuring the resulting oleosin structures. In addition, an oleosin molecule likely interacts with adjacent oleosin molecules for a more stable organization.
The N- and C-terminal portions of an oleosin on the surface of a seed OB may act during germination as a receptor for the binding of lipase for TAG degradation, as a site for ubiquitination for oleosin (and thus PL and TAG) degradation, and/or as a location for interaction with glyoxysomes during germination (Fig. 3). These possibilities can be explored.
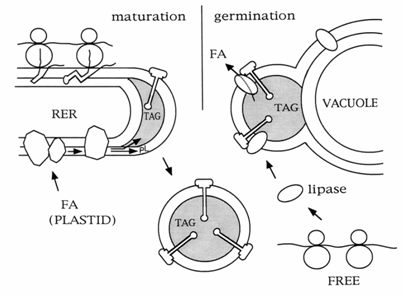
Figure 3. A model of the synthesis and degradation of an oil body in a maize embryo during seed maturation and postgermination. RER, rough endoplasmic reticulum; FA, fatty acid; TAG, triacylglycerols; FREE, free polyribosomes.
3. Synthesis of Oils, Oleosins and Oil Bodies in Endoplasmic Reticulum
Oil bodies, including their constituent TAGs, PLs and oleosins, are synthesized on ER (Fig. 3; [11]). Also, enzymes for the synthesis of TAGs from sn-glycerol-3-phosphate are associated with rough ER (RER) [1]. TAGs synthesized in the ER are sequestered in the hydrophobic region (i.e. the acyl region) of the PL bilayer. Continuation of TAG accumulation at a region of ER forms a budding OB enclosed by a single layer of PLs. This budding OB is stabilized by inclusion of oleosins on its surface.
Despite lacking an N-terminal ER targeting sequence, the nascent oleosin polypeptide associated with a ribosome-mRNA is guided to the ER via the signal-recognition particle pathway [12,13]. The long hydrophobic stretch, and not the N- and C-terminal portions, of oleosin is important in this ER targeting. No specific signal sequence in the hydrophobic stretch is required. Instead, any of the multiple and probably overlapping sequences along the hydrophobic stretch can target the protein to ER. Significantly, the highly conserved proline knot is not important in in vitro targeting of oleosin to microsomes, because replacing the three proline residues with leucine residues does not affect the targeting. Whether this situation occurs in vivo remains to be elucidated. The nascent oleosin polypeptide synthesized or being synthesized on ER assumes a topology based on its hydrophobic and hydrophilic interactions with the PL bilayer. The hydrophilic or amphipathic N- and C-terminal portions interact with the PL layers on the cytosolic side of ER (Fig. 3). The central hydrophobic stretch buries itself in the hydrophobic acyl portion of the PL bilayer. The secondary structure of the 72-residue hydrophobic stretch in the hydrophobic portion of the PL bilayer is unknown, but likely differs from that in a mature OB. The matrix of a mature OB but not the hydrophobic region of ER provides excess hydrophobic volume for the oleosin hydrophobic stretch to assume its presumably most stable hairpin configuration. The oleosin hydrophobic stretch within the hydrophobic region of ER could assume a bended hairpin structure or an extended structure with or without coiling, running parallel to the PL bilayer (Fig. 3). While the ER is synthesizing oleosins, it also produces massive amounts of TAGs, which are temporarily sequestered in and thus enlarge the hydrophobic region of the PL bilayer. Thus, the hydrophobic region of the PL bilayer may have more room for the hydrophobic stretch of a nascent oleosin than that confined by the length of the two acyl chains.
Both the newly synthesized oleosins and TAGs on extensive regions of ER diffuse to budding OBs [6]. This movement is made possible in accordance with the fluid mosaic model of membrane action and thermodynamic considerations. TAGs and the oleosins will be more stable in the hydrophobic environment of a budding OB. During the OB budding process, a gradient of enrichment of these two components should exist from the point of synthesis to the budding OB.
A budding OB is released from the ER as a solitary oil body (Fig. 3). An early release will generate a smaller OB, and vice versa. The size and shape of an OB are determined in part or completely by the relative amount or rate of synthesis of oils and oleosins. High-oil maize kernels (having a high oil-to-oleosin ratio) generated by breeding have large, spherical OBs, whereas low-oil kernels have small OBs with an irregularly shaped surface [14]. In Arabidopsis mutants with reduced oleosin gene activities, the OBs are larger [15,16]. In cells that do not synthesize oleosins, such as those in fatty mesocarp of fruits, the OBs (lipid globules) become very large (following paragraph).
In fatty mesocarp of fruits such as avocado, oil palm and olive, each cell has only one to several large lipid globules, which occupy the bulk of the cell volume [1]. Few or no oleosins are present on the lipid globules. Mesocarp lipids function to attract animals for seed dispersion, and thus their being in small entities as OBs in seeds offers no functional advantages. Mostly likely, TAGs are synthesized in ER, as those in seeds, but without co-synthesis of oleosins (Fig. 3).
Oleosins contain sufficient internal signals for their targeting to intracellular lipid droplets, regardless of their origin. Maize (a monocot) seed OB oleosin is present in transformed Brassica (a dicot) seed OBs [6]. Seed oleosin is present in the intracellular lipid droplets in transformed yeast [13,17] and mammalian cells [5].
4. Abundant Oleosin-Containing Tapetosomes in Flower Tapetum Cells
Oleosins are abundant in flower tapetum cells [18-20]. They are on the surface of numerous lipid droplets, which are structurally and chemically similar to seed OBs. These lipid droplets associate among themselves and with massive ER-derived vesicles, resulting in larger organelles termed tapetosomes [21,22]. Tapetosomes in the Brassicaceae family, especially Brassica and Arabidopsis species, have been studied intensively; whether they are present in other plant families remains to be elucidated.
The tapetum is a single-celled layer enclosing the anther locule, in which microspores mature to become pollen [6]. Tapetum cells are the only anther sporophytic cells that are metabolically very active and control maturation of microspores. At an early stage of anther development, the tapetum cells are specialized for active secretion and contain abundant RER and secretory vesicles. They secrete (a) glucanase to digest the tetrad microspore wall to release solitary microspores, (b) a different glucanase to digest the tapetum wall facing the locule, (c) pollen outer wall (exine) ingredients and (d) likely other materials. At a late stage of anther development, at least in Brassicaceae species, the cells have become chiefly a temporary storehouse of ingredients to be deposited onto maturing pollen as a pollen coat. They are packed with two predominant storage organelles, the elaioplasts and tapetosomes (Fig. 4). The elaioplasts, 3-4 μm in diameter, are specialized plastids largely devoid of thylakoids but filled with small spherical lipid droplets of steryl esters enclosed by the structural protein PAP. Although elaioplasts of similar morphology can be found in non-tapetum cells, such as fruit and petal cells, tapetosomes are unique to the tapetum cells. Each spherical tapetosome, of 2-3 μm in diameter, has oleosin-coated alkane (and some TAG) droplets associated with flavonoid-containing vesicles derived from ER.
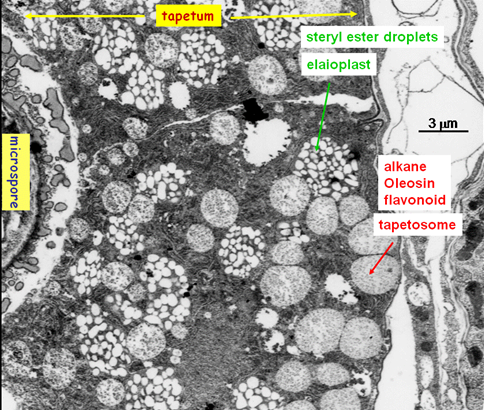
Figure 4. Cross section of a portion of a Brassica anther. The electron micrograph shows the innermost anther-wall cell layer (the tapetum) and a small portion of a microspore in the locule. The tapetum cell is packed with two abundant organelles. The tapetosomes contain alkane, oleosin and flavonoids, and the elaioplasts possess steryl esters. These temporary storage materials will be deposited onto the microspore wall, forming the pollen coat.
The contents of elaioplasts and tapetosomes are discharged to the anther locule after death of the tapetum cells during the final stage of anther development [20-22]. Steryl esters (but not the enclosing structural protein PAP) of elaioplasts, as well as alkanes, oleosins and flavonoids of the tapetosomes (and possibly self-incompatibility protein factors), are transferred to the pollen surface, forming the bulk of pollen coat.
Tapetosomes have a unique morphology [20-22]. Transmission electron microscopy has revealed that in situ each tapetosome has a nonhomogeneous interior possessing tubular structures, which could represent ER-derived vesicles (Fig. 5). The structures of a tapetosome can be observed clearly after it has been isolated and subjected to osmotic swelling. A tapetosome has oleosin-coated alkane droplets associated with ER-derived vesicles via ionic linkage (Fig. 5). Isolated tapetosomes after treatment with a high- or low-pH buffer can be subfractionated into lipid droplets containing alkanes coated with oleosins, and ER-derived vesicles possessing flavonoids.
Figure 5. A model of the synthesis of a tapetosome in a Brassica tapetum cell. (A) Formation of an oleosin-coated lipid droplet from RER with a mechanism similar to that shown in Figure 3. Each lipid droplet consists of an alkane matrix (in blue) enclosed by a layer of phospholipids (red) and oleosins (yellow). (B) Association of several budding oil droplets and their attached ER cisternae. (C) A maturing tapetosome containing attached and detached ER vesicles. (D) A mature tapetosome. (E) Electron micrographs of a tapetosome in situ (left) and an isolated tapetosome after swelling (right).
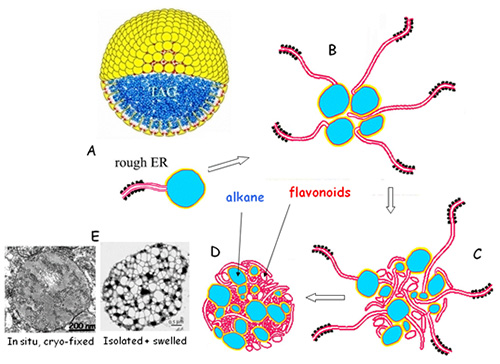
Tapetosomes are synthesized via a special mechanism [20,21]. During early development of a tapetum cell, the ER network contains the luminal protein calreticulin but no oleosins, as revealed by immunofluorescence microscopy. Subsequently, the massive ER network includes both calreticulin and oleosins, and at numerous centers (nascent tapetosomes) of the network, the ratio of oleosin/calreticulin is high. Finally, the ER network largely disappears, and solitary tapetosomes containing oleosins and calreticulin remain. These and other microscopy observations, along with findings from subfractionation of the organelles, allow for construction of a model depicting the biogenesis of tapetosomes from RER (Fig. 5). Initially, alkane droplets are produced via an ER budding mechanism identical to that in maturing seeds. These alkane droplets are covered by oleosins and PLs. Many of these droplets are produced and converge. More ER cisternae are connected to the droplet clusters and eventually break off as vesicles. As a consequence, a tapetosome is produced. During the peak of tapetosome formation, all the maturing tapetosomes in the cell are interconnected via ER cisternae.
5. Characteristics and Functions of the Tapetosome and Pollen-Coat Oleosins
Upon death of the tapetum, the major ingredients of the elaioplasts and tapetosomes are discharged to the pollen surface, forming the pollen coat [20-22]. On the pollen, steryl esters and alkanes form a waterproofing layer, whereas flavonoids protect the haploid pollen from UV radiation and may also serve as signal molecules. The function of the abundant oleosins is less clear.
In Arabidopsis, 9 genes specifically encode the tapetum oleosins, 8 of which are in tandem on chromosome 5, as a result of recent and rapid evolutionary changes [10]. One of these genes is highly expressed to produce an oleosin of 53 kDa, which represents about 70% of all tapetum oleosins [10]. Most of the other Arabidopsis tapetum oleosins are smaller (10-23 kDa). Brassica has a similar tapetum oleosin gene system, and the most active gene (ortholog of the Arabidopsis gene encoding the 53-kDa oleosin) produces a major oleosin of 45 or 48 kDa (from the AA or CC genomes, respectively) [20].
The most abundant pollen-coat oleosin is large (53-kDa in Arabidopsis and 48 or 45 kDa in Brassica) in comparison with most other oleosins (15-25 kDa) because it possesses numerous repeats of short peptides at its C-terminus. Each of these repeats possesses several glycine residues, which makes the protein glycine-rich. Because of its glycine-rich nature, the pollen-coat oleosin (and extrapolating to other oleosins) was initially speculated to interact with the cell walls of the stigma cells. This speculation probably is invalid. Oleosins have undergone rapid evolutionary changes [23,24], and both tapetum and seed oleosins have repeats of short peptides at the C-termini; some of these repeats have high glycine contents, whereas others do not. The rapidity and extensiveness of changes at the C-termini may reflect the minimal structural constraints on this part of the protein to perform functions. The high glycine contents at C-termini of oleosins may be fortuitous, and certainly seed oleosins possessing glycine-rich C-termini do not have an apparent function for interaction with the cell wall. In fact, the short repeats at C-termini of the most abundant tapetum oleosins have a high glycine content and also high serine and lysine contents; so, the oleosin is also serine-rich and lysine-rich (the Arabidopsis 53-kDa oleosin has 26, 16, and 14 mole %, and the Brassica 48-kDa oleosin 21, 16, and 11 mole % of glycine, serine and lysine, respectively).
In addition to stabilizing the alkane droplets in the tapetum cells, oleosins may carry out functions on pollen and subsequently on the stigma because of their amphipathic nature. The N- and C-terminal portions are hydrophilic or amphipathic, and the central portion is hydrophobic. The overall amphipathic oleosin can act as an emulsifying agent to coat uniformly the pollen with steryl esters, alkanes, flavonoids and other ingredients. It can also aid water uptake for germination after the pollen grain has landed on the stigma. Brassicaceae species have dry stigmas, and water must be drawn from the stigma interior to the pollen for germination and tube growth. Steryl esters and alkanes are not amphipathic and cannot act as a wick. The abundant and amphipathic oleosins (and flavonoids) could be such a wick. On the basis of these proposed functions, the mutational addition of repeats of short peptides, which are all fairly hydrophilic, to the C-termini and the observed in vivo fragmentation of the Brassica 45/48-kDa oleosins into two halves (probably fortuitous) do not affect functioning of the oleosins. The proposed functions are in agreement with the observation that the pollen of an Arabidopsis mutant null in the major pollen-coat oleosin does not hydrate efficiently on the stigma [25]. This partial loss of function could have been due to the lack of sufficient oleosins in stabilizing the alkane droplets in the tapetum cells, emulsifying properly the lipids and other ingredients of the pollen coat, and/or acting as a wick of the pollen-coat for water uptake on the stigma.
Overall, the major structural features in oleosins for performing the proposed functions are a long hydrophobic hairpin to interact with the alkane droplets in tapetosomes and an amphipathic molecule to emulsify the pollen coat materials and take up water from stigma. All the observed mutational changes on tapetum oleosins have not affected these essential features and are thus tolerated because of the lack of sufficient selective pressure.
6. Evolution of Triacylglycerols, Oil Bodies, Oleosins and Tapetosomes
Prokaryotes, in general, do not store TAGs as food reserves. A minor exception occurs in Actinomyces, which produce TAGs under certain nutritional and other environmental conditions. TAGs likely evolved as efficient food reserves in a primitive eukaryote by the addition of one enzyme, diacylglycerol acyltransferase, which evolved from one of the existing acyltransferases. Diacylglycerol acyltransferase diverted DAGs from the ubiquitous PL metabolic pathway to TAGs. Initially, the hydrophobic TAGs were present between the two PL layers of the ER membrane, where diacylglycerol acyltransferase was located. Today, seeds of certain species still have some TAGs present along the hydrophobic region of the PL bilayers in ER [1].
The presence of excess TAGs in the ER membrane would interfere with the normal functioning of the ER. Removal of the TAGs from ER via budding to become solitary droplets would offer a functional advantage. The droplets, each containing a TAG matrix enclosed by a layer of PLs originating from ER, would be unstable. In yeasts, the droplets were made more stable with a coat of amphipathic proteins, especially the TAG- and steryl ester-synthesizing and hydrolyzing enzymes, and the semistability would enable the droplets to undergo dynamic metabolic fluxes. In mammals, the droplets evolved to different forms with proteins and membranes, and they are semistable and yet amenable to easy metabolic fluxes. In plants, the droplets were stabilized by the evolutionary appearance of oleosins, and so they were of small size and amenable to prolonged storage in desiccated seeds and yet quick metabolic mobilization during germination. Oleosins and their coated oil droplets have been found in diploid and triploid storage sporophytic cells of seeds of angiosperms and gymnosperms, haploid storage cells of female gametophytes (in gymnosperm seeds), haploid cells of male gametophytes (pollen), the moss Physcomitrella (in both diploid sporophyte and haploid gametophyte) and diploid sporophytic cells of floral tapeta.
The hydrophobic stretch of 72 resides in oleosins is the longest, and is actually more than twice as long as any found in all prokaryotic and eukaryotic proteins. The mechanism by which the stretch evolved is intriguing. A hypothesis has been proposed on the basis of the following observations: (A) the length of 72 nonpolar residues is about four times that of a transmembrane polypeptide (~20 residues); (B) several relatively polar residues are present along both the antiparallel stretches; and (C) a certain degree of residue symmetry exists along the two antiparallel stretches. The hypothesis suggests that the long hydrophobic stretch evolved from duplications or gradual extension of a transmembrane peptide of an ER protein in a primitive eukaryote. The hairpin hydrophobic stretch of oleosin is conserved, but the N- and C-terminal portions have undergone extensive evolutionary changes because of the limited structural and functional constraints.
Oleosin-coated oil droplets in diverse plant species can be categorized into two groups according to their structures and functions. The solitary oleosin-coated oil bodies in seeds and pollen store TAGs for germination and postgerminative growth in the respective organs. The tapetosomes contain clustered oleosin-coated alkane droplets associated with ER-derived flavonoid-containing vesicles and store and deliver materials to the surface of maturing pollen. Whether, during evolution, solitary oleosin-coated alkane droplets similar to the modern seed oil bodies appeared before the complex tapetosomes, or vice versa, can be speculated. The most primitive plant known to contain oleosin is the moss Physcomitrella. The Physcomitrella oleosin is associated with storage oil bodies in the sporophyte and gametophyte. This moss does not have flowers or tapetum and thus would not have analogs of tapetosomes. Brassicaceae species contain abundant tapetosomes in tapeta, but maize and rice tapeta do not have any. Thus, tapetosomes likely evolved from solitary oleosin-coated lipid droplets similar to the modern oil bodies in seeds. Initially, these ancestor droplets, in the tapetum, solitary or in groups, delivered lipids and oleosins to the pollen surface. Subsequently, they became associated with vesicles that contain flavonoids and other materials for the pollen surface. Thus, tapetosomes evolved to perform the overall function of packaging and storing materials for delivery to the pollen surface.
Chlamydomonas and Volvox genomes have genes that could encode oleosin-like proteins. These putative proteins have the 11-residue hairpin loop (PX5SPX3P, X being a nonpolar residue) that is highly conserved among oleosins of diverse plant species. However, the nonpolar sequences adjacent to the hairpin loop are short, and the length of the whole nonpolar sequence is similar to that of a transmembrane segment in diverse proteins. Whether these putative oleosin-like proteins are present on the OBs of the algae and if so what the percentages are of the protein being on the OBs and membrane (e.g. of the ER) remain to be elucidated.
References
- Huang, A.H.C. Oil bodies and oleosins in seeds. Annu. Rev. Plant Physiol. Mol. Biol., 43, 177-200 (1992).
- Napier, J.A., Stobart, A.K. and Shewry, P.R. The structure and biogenesis of plant oil bodies: the role of the ER membrane and the oleosin class of proteins. Plant Mol. Biol., 31, 945-956 (1996).
- Galili, G., Sengupta-Gopalan, C. and Ceriotti, A. The endoplasmic reticulum of plant cells and its role in protein maturation and biogenesis of oil bodies. Plant Mol. Biol., 38, 1-29 (1998).
- Frandsen, G.I., Mundy, J. and Tzen, J.T. Oil bodies and their associated proteins, oleosin and caleosin. Physiol. Plant., 112, 301-307 (2001).
- Murphy, D.J. The biogenesis and functions of lipid bodies in animals, plants and microorganisms. Plant J., 13, 1-16 (2001).
- Hsieh, K. and Huang, A.H.C. Endoplasmic reticulum, oleosins, and oils in seeds and tapetum. Plant Physiol., 136, 3427-3434 (2004) (DOI: 10.1104/pp.104.051060).
- Bickel, P.E., Tansey, J.T. and Welte, M.A. PAT proteins, an ancient family of lipid droplet proteins that regulate cellular lipid stores. Biochim. Biophys. Acta, 1791, 419-440 (2009) (DOI: 10.1016/j.bbalip.2009.04.002).
- Beller, M., Thiel, K., Thul, P.J. and Jackle, H. Lipid droplets: a dynamic organelle moves into focus. FEBS Lett., 584, 2176-2182 (2010) (DOI: 10.1016/j.febslet.2010.03.022).
- Huang, C.Y., Chung, C.I., Lin, Y.C., Hsing, Y.I.C. and Huang, A.H.C. Oil bodies and oleosins in Physcomitrella possess characteristics representative of early trends in evolution. Plant Physiol., 150, 1192-1203 (2009) (DOI: 10.1104/pp.109.138123).
- Kim, H.U., Hsieh, K., Ratnayake, C. and Huang, A.H.C. A novel group of oleosins is present inside the pollen of Arabidopsis. J. Biol. Chem., 277, 22677-22684 (2002) (DOI: 10.1074/jbc.M109298200).
- Qu, R. , Wang, S.M., Lin, Y.H., Vance, V.B. and Huang, A.H.C. Characterization and biosynthesis of membrane proteins of lipid bodies in the scutella of maize (Zea mays L.). Biochem. J., 235, 57-65 (1986).
- Abell, B.M., Hahn, M., Holbrook, L.A. and Moloney, M.M. Membrane topology and sequence requirements for oil body targeting of oleosin. Plant J., 37, 461-470 (2004).
- Beaudoin, F. and Napier, J.A. Targeting and membrane-insertion of a sunflower oleosin in vitro and in Saccharomyces cerevisiae: the central hydrophobic domain contains more than one signal sequence, and directs oleosin insertion into the endoplasmic reticulum membrane using a signal anchor sequence mechanism. Planta, 215, 293-303 (2002).
- Ting, J.T.L., Lee, K., Ratnayake, C., Platt, K.A., Balsamo, R.A. and Huang, A.H.C. Oleosin genes in maize kernels having diverse oil contents are constitutively expressed independent of oil contents: size and shape of intracellular oil bodies are determined by the oleosin/oils ratio. Planta, 199, 158-165 (1996).
- Siloto, R.M.P., Findlay, K., Lopez-Villalobos, A., Yeung, E.C., Nykiforuk, C.L. and Moloney, M.M. The accumulation of oleosins determines the size of seed oilbodies in Arabidopsis. Plant Cell, 18, 1961-1974 (2006) (DOI: 10.1105/tpc.106.041269).
- Shimada, T.L., Shimada, T., Takahashi, H., Fukao, Y. and Hara-Nishimura, I. A novel role for oleosins in freezing tolerance of oilseeds in Arabidopsis thaliana. Plant J., 55, 798-809 (2008) (DOI: 10.1111/j.1365-313X.2008.03553.x).
- Ting, J.T.L., Balsamo, R.A., Ratnayake, C. and Huang, A.H.C. Oleosin of plant seed oil bodies is correctly targeted to the lipid bodies in transformed yeast. J. Biol. Chem., 272, 3699-3706 (1997).
- de Oliveira, D.E., Franco, L.O., Simoens, C., Seurinck, J., Coppieters, J., Botterman, J. and Van Montagu, M. Inflorescence-specific genes from Arabidopsis thaliana encoding glycine-rich proteins. Plant J., 3, 495-507 (1993).
- Roberts, L.S,, Gerster, J., Allard, S., Cass, L. and Simmonds, J. Molecular characterization of two Brassica napus genes related to oleosins which are highly expressed in the tapetum. Plant J., 6, 927-933 (1994).
- Wu, S.S.H., Platt, K.A., Ratnayake, C., Wang, T.W., Ting, J.T.L. and Huang, A.H.C. Isolation and characterization of novel neutral-lipid-containing organelles and globuli-filled plastids from Brassica napus tapetum. Proc. Natl. Acad. Sci. U.S.A., 94, 12711-12716 (1997).
- Hsieh, K. and Huang, A.H.C. Lipid-rich tapetosomes in Brassica tapetum are composed of oleosin-coated oil droplets and vesicles, both assembled in and then detached from the endoplasmic reticulum. Plant J., 43, 889-899 (2005) (DOI: 10.1111/j.1365-313X.2005.02502.x).
- Hsieh, K. and Huang, A.H.C. Tapetosomes in Brassica tapetum accumulate endoplasmic reticulum-derived flavonoids and alkanes for delivery to the pollen surface. Plant Cell, 19, 582-596 (2007).
- Schein, M., Yang, Z.H., Mitchell-Olds, T. and Schmid, K.J. Rapid evolution of a pollen-specific oleosin-like gene family from Arabidopsis thaliana and closely related species. Mol. Biol. Evol., 21, 659-669 (2004).
- Fiebig, A., Kimport, R. and Preuss, D. Comparisons of pollen coat genes across Brassicaceae species reveal rapid evolution by repeat expansion and diversification. Proc. Natl. Acad. Sci. U.S.A., 101, 3286-3291 (2004).
- Mayfield, J.A. and Preuss, D. Rapid initiation of Arabidopsis pollination requires the oleosin-domain protein GRP17. Nature Cell Biol., 2, 128-130 (2000).
Acknowledgements
The research was supported by the National Science Foundation (MCB-0131358), the U.S. Department of Agriculture (National Research Initiative Competitive Grant No. 2004-02429) and research grants from the Academia Sinica and the National Research Council of Taiwan.
In This Section
- Plant Fatty Acid Synthesis
- Production of Unusual Fatty Acids in Plants
- Arabidopsis Acyl-Coenzyme A-Binding Proteins
- Long Chain acyl-coA Synthetases and Other Acyl Activating Enzymes
- Plant Triacylglycerol Synthesis
- Triacylglycerol Biosynthesis in Eukaryotic Microalgae
- Subcellular Oil Droplets and Oleosins in Plants
- Triacylglycerol Mobilisation in Plants
- Role of Transcription Factors in Storage Lipid Accumulation in Plants
- Biosynthesis of Plant Lipid Polyesters
- Rubber Biosynthesis in Plants
- Carotenoid Biosynthesis and Regulation in Plants
- The Oxylipin Biosynthetic Pathways in Plants
- N-Acylphosphatidylethanolamines (NAPEs), N-acylethanolamines (NAEs) and Other Acylamides: Metabolism, Occurrence and Functions in Plants
- Phosphoinositide Signaling in Plants
- Plant Lipidomics
- 50 years of Galactolipid Research: The Beginnings
- Transport and function of lipids in the plant phloem