Plant Fatty Acid Synthesis
The Author: John L. Harwood, School of Biosciences, Cardiff University, Cardiff CF10 3AX, Wales, U.K.
Introduction
Plants synthesise a huge variety of fatty acids although only a few are major and common constituents [1]. Broadly speaking, long-chain fatty acids are synthesised de novo from small precursors ultimately derived from photosynthate. Two enzyme systems are utilised, acetyl-CoA carboxylase and fatty acid synthase (Fig. 1). The end products of this synthesis are usually the saturated fatty acids palmitate and stearate with the latter predominating (in most plants by 2-3 times that of palmitate). Once the long-chain acids have been produced they can be subject to elongation, desaturation and further modifications (Fig. 1). Unlike acetyl-CoA carboxylase and fatty acid synthase, which are soluble enzymes, the elongases are membrane-bound and sited in the endoplasmic reticulum. Only recently have the details of such reactions started to be elucidated at the molecular level. Elongases are coded by FAE genes while the desaturases are coded by FAD genes.
Figure 1. Fatty acid biosynthesis in plants.
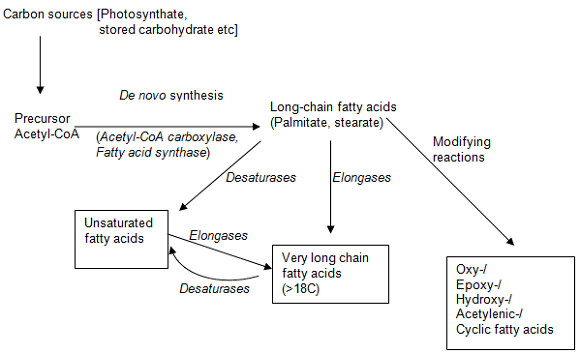
Fatty acid desaturases are usually membrane-bound and utilise complex lipid substrates such as phosphatidylcholine or monogalactosyldiacylglycerol [2]. An exception is the stearoyl-acyl carrier protein (ACP) Δ9-desaturase that is present in the chloroplast stroma and converts stearate to oleate. When desaturases produce polyenoic fatty acids, the latter usually have a methylene-interrupted structure, such as linoleic (cis, cis Δ9,12-octadecadienoic) acid or α-linolenic (all cis Δ9,12,15-octadecatrienoic) acid.
Some plants can produce unusual fatty acids in their seed oils, many of which have useful industrial applications [1,3,4]. These include hydroxyl fatty acids, cyclopropane fatty acids, epoxy fatty acids and conjugated unsaturated fatty acids. It is noteworthy that these unusual fatty acids accumulate preferentially in triacylglycerols and are essentially excluded from membrane acyl lipids – presumably because they would impair function.
De novo Synthesis
Source of carbon
Apart from germination, photosynthate provides the source of carbon for de novo fatty acid synthesis. Early work highlighted a controversy as to how much carbon could be derived directly from plastid pyruvate dehydrogenase [5]. More recent studies [see 6] have concluded that this enzyme provides most of the acetyl-CoA needed for leaf fatty acid biosynthesis. In contrast, the situation maybe more complex in developing oil seeds such as Brassica napus (oilseed rape) [see 6].
Acetyl-CoA carboxylase
The first committed step in fatty acid biosynthesis is catalysed by acetyl-CoA carboxylase (EC 6.4.1.2). This belongs to the group of soluble Class 1 biotin-containing enzymes which use ATP and bicarbonate to carboxylate a biotin prosthetic group (Fig. 2). The carboxy group is then transferred to the acceptor acetyl-CoA to form malonyl-CoA. The initial partial reaction is catalysed by biotin carboxylase, uses bicarbonate (rather than carbon dioxide) as the source of carbon and acts via a carboxyphosphate intermediate [7]. The second partial reaction is catalysed by carboxyltransferase (Fig. 2). During the reaction, electron transfer is thought to allow direct reaction of carbon dioxide with the incoming acetyl-CoA to yield malonyl-CoA [7]. While the various Class 1 biotin-containing carboxylases share a very similar sequence identity for their biotin carboxylases, the carboxyltransferases are distinct and give the specificity to the overall reaction [6].
Figure 2. Reactions of acetyl-CoA carboxylase.
(A) The biotin prosthetic group which is attached to biotin carboxyl carrier protein (BCCP) via a lysine residue. The carboxylation takes place on a nitrogen in the upper ring (as shown).
(B) The partial reactions of acetyl-CoA carboxylase.
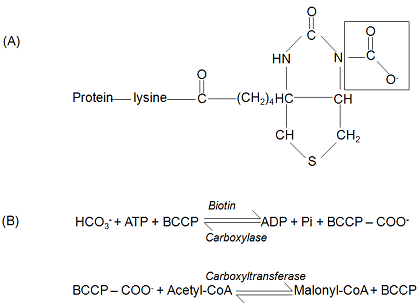
There are two distinct molecular forms of acetyl-CoA carboxylase in plants – multiprotein complexes and multifunctional proteins. Furthermore, plants also have isoforms in two subcellular sites. A plastid-localised isoform is used for de novo synthesis of fatty acids while an extra-plastid isoform (presumed to be cytosolic) provides malonyl-CoA for fatty acid elongation as well as other functions [2,6]. The different susceptibility of grasses and dicotyledons to various herbicides belonging to the aryloxypropionate and cyclohexanedione chemical groups (which both inhibited fatty acid synthesis in grasses) led to the discovery that, while grasses had two different multifunctional protein forms of acetyl-CoA carboxylase (mol. masses of 220-230 kDa), the dicotyledons have a multifunctional protein in the cytosol but a multienzyme complex in the stroma [2,6]. The latter was coded by 4 separate genes. Three of these genes (biotin carboxylase, biotin carboxyl carrier protein, α-subunit of carboxyltransferase) are nuclear-encoded. The fourth gene coding for the β-subunit of carboxyltransferase is plastid-located [6].
For further details of the reaction characteristics, molecular structure, herbicide sensitivity and genetics of acetyl-CoA carboxylase see [6].
Regulation of acetyl-CoA carboxylase
Since acetyl-CoA carboxylase catalyses the first committed reaction in fatty acid synthesis, it might be thought a good candidate for important regulation – as revealed in animal tissues [8]. Two types of experiment confirmed that the regulation of acetyl-CoA carboxylase was important for the control of lipid synthesis – at least in leaf tissues. First, Post-Beittenmiller et al. [9] measured pool sizes of acyl-thioesters during dramatic changes in the rate of fatty acid formation. The data provided experimental evidence pointing to the importance of acetyl-CoA carboxylase in regulating overall synthesis. By making use of the specific action of grass-selective herbicides, Page et al. [10] were able to measure the flux control exerted by the enzyme for lipid synthesis in barley or maize leaves. Their results showed that the acetyl-CoA carboxylase reaction alone controlled about 55% of the total flux, thus demonstrating clearly its important role in regulation.
Fatty acid synthase
The second enzyme complex involved in de novo synthesis is fatty acid synthase (FAS) [2,6]. In plants this is a Type II FAS consisting of a multiprotein complex. There are a number of enzymes involved in FAS (Fig. 3). These are used for acyl-transfer, the four sequential reactions involved in 2-carbon addition and in termination of the overall reaction. In addition, acyl carrier protein (ACP) is used as the acyl carrier for the various intermediate reactions. Although de novo synthesis is located in the stroma, plant mitochondria are capable of limited fatty acid synthesis. This is probably mainly used for lipoic acid formation [see 6]. Nevertheless, three genes for mitochondrial ACP have been detected in Arabidopsis. In addition, this plant has five deduced genes for plastid ACP [6].
Figure 3. The overall reaction of fatty acid synthase.
* The first condensation reaction is catalysed by β-ketoacyl-ACP synthase III (KAS III), which uses acetyl-CoA and malonyl-ACP substrates. The next six condensations are catalysed by KAS I while the final reaction between palmitoyl-ACP and malonyl-ACP utilises KAS II.
Figure taken from reference 8 with permission from the author and the publisher, Blackwell Publishing Ltd.
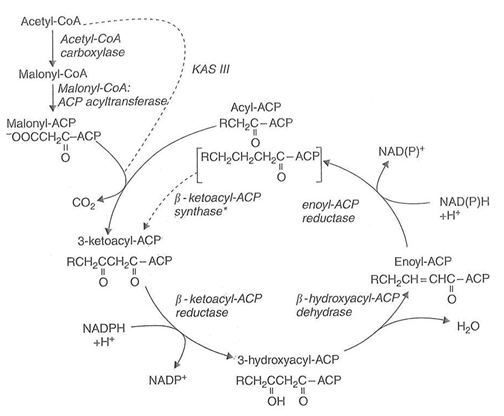
Although acetyl-CoA:ACP acyltransferase has been studied in plants [11], its function has been put in doubt by the clear demonstration of a short-chain condensing enzyme (KAS III) in plants [12,13] (see later). However, the importance of malonyl-CoA:ACP acyltransferase is not in doubt and some details of this enzyme will be found in [2].
For the successive addition of two-carbon units four enzyme reactions are needed. These are catalysed by a condensing enzyme (β-ketoacyl-ACP synthase, KAS), first reductase (β-ketoacyl-ACP reductase), dehydrase (β-hydroxyacyl-ACP dehydrase) and a second reductase (enoyl-ACP reductase) (Fig. 3). Three different condensing enzymes are found generally in plants – called KAS I, II and III. A fourth one may be present in some tissues [6]. The initial condensation of two 2-carbon units is catalysed by KAS III (Table 1) which uses acetyl-CoA and malonyl-ACP substrates. After the two reductions and dehydration reactions a 4-carbon fatty acid, butyrate, is produced. This is poorly condensed by KAS III but is a good substrate for KAS I, which acts to elongate 4- to 14- carbon chains (Table 1). The final condensation is catalysed by KAS II which is mainly responsible for the production of stearate (Table 1). KAS I and KAS II can be distinguished from each other by their differential sensitivity to cerulenin and arsenite, respectively [2,11]. As one might anticipate, the plant FAS and, particularly its condensing enzyme isoforms, has many parallels with the classic Type II FAS from Escheria coli [14].
| Table 1. Comparison of the condensing enzymes of fatty acid synthase | |
| KAS I | Can use 2-14C acyl-ACPs for condensation with malonyl-ACP. Utilised for making 6-16C fatty acids. Cerulenin sensitive. Subunit of 43-46 kDa. Works as homodimer but with variations in barley. Has a cys-his-his triad at its active site. |
| KAS II | Can use palmitoyl-ACP > myristoyl-ACP for condensation with malonyl-ACP. Used for chain lengthening of palmitate to stearate and, hence, controls the final ratio of 16C/18C products. Native enzyme has molecular mass of 88 kDa. Has cys-his-his triad at active site. Arsenite sensitive but poorly inhibited by cerulenin. |
| KAS III | Responsible for initial condensation reaction. Uses acetyl-CoA and malonyl-ACP. Thiolactomycin sensitive. Cerulenin insensitive. Subunit mol. mass 37-40 kDa. Exists as homodimer. Has cys-his-asn triad at active site. |
Details of the molecular structure and genetics of the individual condensing enzymes will be found in [6]. Moreover, there are also some KAS enzymes with unusual properties which are able to cope with particular fatty acid substrates (unsaturated or very-long-chain) in specific tissues [6]. The crystal structure of E. coli KAS III has been deduced [15] and allowed, by sequence comparison, some speculation as to the active site of the plant KAS III [16]. This showed that key site residues (Cys-His-Asn triad) were completely conserved and allowed preferred conformations of other parts of the adjacent protein structure to be deduced [16]. Although it has been suggested that condensing enzymes may be important for regulating carbon flux, experiments to demonstrate this have not been convincing [6].
After condensation, the intermediate is reduced by β-ketoacyl-ACP reductase (Fig. 3). Usually this is regarded as a NADPH-utilising enzyme. It has been purified from Brassica napus (subunit mol. mass 28 kDa) and functions as a tetramer [17]. The reductase from oilseed rape has also been crystallised and features of its substrate binding and catalysis deduced [see 6]. At certain developmental stages, this reductase may exert significant influence on the rate of oil accumulation in Brassica [6].
The third enzyme in the elongation cycle is β-hydroxyacyl-ACP dehydrase, which has been purified from spinach leaves. It is encoded by two genes in Arabidopsis and is specific for the D(-)substrate stereoisomer.
The fourth enzyme is enoyl-ACP reductase (Fig. 3). Depending on the plant source, the reductase may be specific for NADH or may be able to utilise NADPH as well. The reductase from Brassica has a subunit molecular mass of 35 kDa. Quite a lot is known about the gene (or genes) coding for the enoyl reductase, as well as the reaction mechanism which involves a compulsory-ordered ternary complex [18].
Enoyl reductase is a target for a metabolite of isoniazid, which is used for the treatment of tuberculosis. In addition, the commonly used antibiotic, triclosan, targets the enzyme both in E. coli and plants [19]. Some carotenoid synthesis herbicides may inhibit the enzyme as a secondary mode of action [see 6].
Termination mechanisms
In plants the process of de novo fatty acid synthesis can be halted in various ways. For complex lipid synthesis within the plastid (especially the phosphatidylglycerol component of the thylakoid membranes), acyl-ACPs such as palmitoyl-ACP or stearoyl-ACP, can be used directly by acyltransferases of the Kornberg-Pricer pathway producing phosphatidate. This usage is favoured because β-ketoacyl-ACP synthase II (KAS II) uses myristoyl-ACP and palmitoyl-ACP (but not stearoyl-ACP) preferentially as substrates [2,11].
Alternatively, the acyl-ACP products of fatty acid synthase can be hydrolysed by thioesterase(s). The release of unesterified fatty acids allows them to be exported outside the plastid to undergo modifications on the endoplasmic reticulum (see later) or to be used for complex lipid biosynthesis in the extra-plastidic compartment. Fatty acids which are exported take part in the ‘eukaryotic’ pathway of lipid synthesis whereas fatty acids retained in the plastid are used for the ‘prokaryotic’ pathway [20]. Lipids made by the former pathway characteristically are enriched at both the sn-1 and sn-2 positions with 16C fatty acids whereas the prokaryotic pathway produces thylakoid lipid molecules with 18C acids at the sn-2 position.
Two classes of fatty acyl-ACP thioesterases have been described (FATA and FATB). FATA preferentially hydrolyses oleoyl-ACP whereas FATB has highest activity with saturated acyl-ACPs. In Arabidopsis there are two genes for FATA and one for FATB. Although most plants contain FATB enzymes that have good activity with substrates in the 14-18C range, some species have a particular need for thioesterases that can act on shorter chains. Thus, tissues such as coconut, California bay, palm kernel or developing Cuphea seeds produce oils with short or medium acyl chains. The medium-chain FATB from California bay was the first to be cloned and has been used to transform oilseed rape. Seeds from the latter could produce up to 60% of their total fatty acid contents as laurate [21]. Further information on these thioesterases is given in [2,6].
Fatty Acid Desaturases
The introduction of double bonds into the fatty acid chain relies on the activity of desaturases. In plants, such desaturases work via an aerobic mechanism with oxygen being reduced by 4H, two from the substrate fatty acid and two from the reductant used (Fig. 4).
Figure 4. A simplified depiction of the aerobic desaturation mechanism in plants.
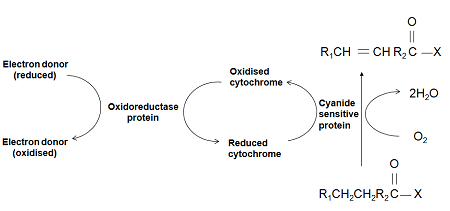
Electron donors, cytochromes and substrates can all vary depending on the desaturase. Examples of donors could be NADH, NADPH or reduced ferridoxin while cytochrome b5 is the usual cytochrome used. Substrates include acyl-ACPs, acyl-CoAs, phosphatidylcholine and monogalactosyldiacylglycerol. Fatty acids can be saturated, monoenoic or polyenoic, and the position of the new double bond varies.
Genes coding for desaturases are abbreviated as FAD or fad. These are numbered generally according to the position of the double bond introduced as well as the nature of the substrate used (Fig. 5) [22].
Figure 5. The two-pathway scheme for membrane glycerolipid synthesis in Arabidopsis leaves (see ref. 2 for details).
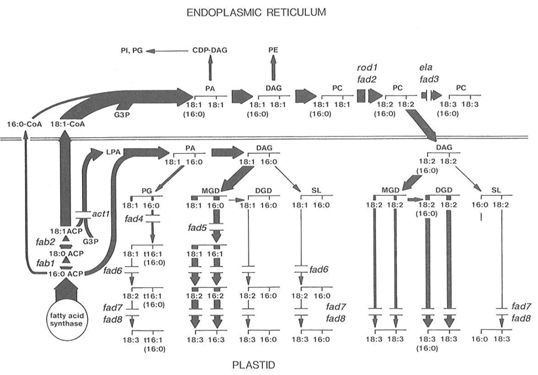
Widths of the lines show the relative fluxes through different reactions. The breaks show the putative enzyme deficiencies in various known mutants.
Figure taken from reference 22 with permission from the author and the publisher, Elsevier.
Stearoyl-ACP Δ9-desaturase
In contrast to other desaturases, this is a soluble enzyme in the plastid stroma that converts stearate into oleate. Its soluble nature has allowed structural studies to be performed [2,23]. Genes for the enzyme have been identified in a whole variety of plants [23,24]. Moreover, in addition to the usual stearoyl/palmitoyl-ACP Δ9-desaturases, there are other soluble acyl-ACP desaturases in different plants. For example, Δ4 and Δ6 palmitoyl-ACP desaturases and a Δ9-myristoyl-ACP have been reported [23,24]. They all have considerable sequence homology which is notably different from acyl-lipid desaturases or to the acyl-CoA desaturase of mammals, yeast or the red alga C. merolae [24].
The soluble Δ9-desaturases can be engineered with amino acid substitutions or chimeric proteins produced which have novel properties [see 24].
Δ12-Desaturase
The most abundant plant fatty acids are linoleic and α-linolenic acids and these are produced by further desaturation of oleate with the introduction of methylene-interrupted double bond arrangements (Fig. 6). Earlier work on the Δ12-desaturase enzyme producing linoleate has been summarised [11]. Complex lipids are substrates and the desaturation can take place on the endoplasmic reticulum (FAD 2 on phosphatidylcholine) or within the plastid (FAD 6 on glycosylglycerides) (Fig. 5). For seed oils FAD 2 is the main pathway while in leaves the eukaryotic pathway utilising FAD 2 will operate to varying extents [22]. In most species of higher plants phosphatidylglycerol is the only exclusive product of the prokaryotic pathway and the other thylakoid lipids are produced by the eukaryotic pathway. In other species such as spinach or Arabidopsis, these latter are made equally by the two pathways [22].
Figure 6. The sequential desaturation of stearate to α-linolenate.
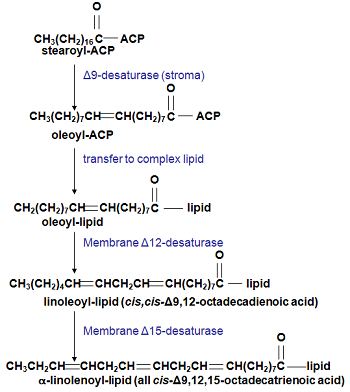
Stearoyl-ACP produced by fatty acid synthase is the substrate for the Δ9-desaturase (also in the stroma) which forms oleoyl-ACP. In the prokaryotic pathway, oleate is then incorporated into chloroplast membrane lipids for further desaturation at the Δ12-(FAD 6 enzyme) and Δ15-(FAD 7, FAD 8 enzymes) positions (see Fig. 5). For the eukaryotic pathway, oleoyl-ACP is hydrolysed by FATA and/or FATB and the unesterified oleate used to form oleoyl-CoA by acyl-CoA synthase on the plastid envelope [2]. The oleoyl-CoA can then be incorporated into phosphatidylcholine by various reactions where it forms a substrate for the Δ12-desaturase (FAD 2) to form linoleate and then a Δ15-desaturase (FAD 3) to yield α-linolenate. As shown in Figure 5 the diacylglycerol from phosphatidylcholine can be released to be incorporated into chloroplast lipids for further desaturation by FAD 7/8.
Production of α-linolenate
The world’s most prevalent fatty acid is α-linolenate. The desaturation of linoleate to form α-linolenate (Fig. 6) uses a Δ15-desaturase (or omega-3 desaturase?) acting on linoleate acylated to a complex lipid. While FAD 3 uses phosphatidylcholine, FAD 7 and FAD 8 use chloroplast glycosylglycerides. Thus, in most leaves the formation of this major metabolic product uses a complex series of enzyme reactions in several distinct locations in the cell (see Fig. 6). As befits a membrane-located enzyme, we know rather little about the detailed structure and mechanism of the Δ15-desaturase [2].
Other membrane desaturases
Two other chloroplast desaturases, FAD 4 and FAD 5, act on complex lipid substrates. Both are highly substrate-selective in contrast to the other acyl-lipid desaturases [22]. The FAD4 enzyme inserts a trans-Δ3 double bond into palmitate at the sn-2 position of phosphatidylglycerol. It is responsible for the significant amount of this most unusual plant fatty acid in phosphatidylglycerol which has functions in photosynthesis [25] and consequences for low temperature susceptibility [26].
FAD 5 is responsible for the synthesis of Δ7-hexadecenoate on monogalactosyldiacylglycerol and, possibly, digalactosyldiacylglycerol [22].
A few plants (such as evening primrose or borage) contain a Δ6-desaturase. Such enzymes contain the cytochrome donor component fused to the desaturase protein. Because the Δ6-desaturase acts between an existing double bond and the carboxyl end of the acid it has been termed a ‘front-end desaturase’ [27].
Variants on the desaturase theme
As mentioned above, plants can contain a remarkable variety of fatty acids [1], many of which have important (or potential) industrial uses [28-31]. These include hydroxylases, conjugases, acetylenases, oxygenases and enzymes making trans-desaturated products [24]. There are also cyclopropane or cyclopropene-ring acids [1] though rather little is known of their formation.
Recently, with the renewed awareness of the nutritional importance of very-long-chain n-3 polyunsaturated fatty acids [32] and the potential supply problems from (over)fishing, there have been determined attempts to engineer crops for suitable oils [27]. This has met with some success but more work is clearly needed before commercially successful crops can be grown. In contrast, some algae (and bacteria) can produce such acids (eicosapentaenoic (EPA) and docosahexaenoic (DHA) acids) using a polyketide synthase metabolic route [33].
For further details of the production of unusual plant fatty acids see [1,23,24,27-31] and the 'A Lipid Primer' webpages.
Fatty acid elongation
Elongation of long-chain fatty acids to produce very-long-chain (>18C) products (both saturated and unsaturated) requires the sequential use of 4 reactions to add each 2C unit [2]. These reactions are analogous to de novo synthesis (i.e. condensation, 1st reduction, dehydration, 2nd reduction) but there are several differences. Notably these elongases are membrane-bound enzymes which use acyl-CoA substrates and malonyl-CoA directly as the source of the 2C unit. Earlier work is summarised in [2,34] and some more recent findings will be found in [27,35].
References
- Gunstone, F.D., Harwood, J.L. and Dijkstra, A.J. (eds.) The Lipid Handbook (Third Edition) (Taylor & Francis, Boca Raton, Florida), (2007).
- Harwood, J.L. Recent advances in the biosynthesis of plant fatty acids. Biochim. Biophys. Acta, 1301, 7-56 (1996).
- Kinney, A.J. Perspectives on the production of industrial oils in genetically engineered oilseeds. In: Lipid Biotechnology, pp. 85-93 (T.M. Kuo and H.W. Gardner (eds.), Marcel Dekker, New York) (2001).
- Broun, P., Gettner, S. and Somerville, C. Genetic engineering of plant lipids. Annu. Rev. Nutr., 19, 197-216 (1999).
- Leidvogel, B. Lipid precursors in plant cells: the problem of acetyl-CoA generation for plastid fatty acid synthesis. In: The Metabolism, Structure and Function of Plant Lipids, pp. 509-512 (P.K. Stumpf, J.B. Mudd and W.D. Nes (eds.), Plenum, New York) (1987).
- Harwood, J.L. Fatty acid biosynthesis. In: Plant Lipids: Biology, Utilisation and Manipulation, pp. 27-66 (D.J. Murphy (ed.), Blackwell Publishing, Oxford) (2005).
- Knowles, J R. The mechanism of biotin-dependent enzymes. Annu. Rev. Biochem., 58, 195-221 (1989).
- Gurr, M.I., Harwood, J.L. and Frayn, K.N. Lipid Biochemistry (Fifth Edition) (Blackwell Publishing, Oxford) (2002).
- Post-Beittenmiller, D., Roughan, P.G. and Ohlrogge, J.B. Regulation of plant fatty acid biosynthesis. Analysis of acyl-CoA and acyl-ACP substrate pools in spinach and pea chloroplasts. Plant Physiol., 100, 923-930 (1992).
- Page, R.A., Okada, S. and Harwood, J.L. Acetyl-CoA carboxylase exerts strong flux control over lipid synthesis in plants. Biochim. Biophys. Acta, 1210, 369-372 (1994).
- Harwood, J.L. Fatty acid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol., 39, 101-138 (1988).
- Jaworski, J.G., Clough, R.C. and Barnum, S.R. A cerulenin-insensitive short-chain 3-ketoacyl acyl carrier protein synthase in Spinacea oleraceae. Plant Physiol., 90, 41-44 (1989).
- Walsh, M.C., Klopfenstein, W.E. and Harwood, J.L. The short chain condensing enzyme has a widespread occurrence in the fatty acid synthetases in higher plants. Phytochemistry, 29, 3797-3799 (1990).
- Heath, R.J., White, S.W. and Rock, C.O. Lipid biosynthesis as a target for antibacterial agents. Prog. Lipid Res., 40, 467-497 (2001).
- Qui, X.Y., Janson, C.A., Konstantinides, A.K. et al. Crystal structure of β-ketoacyl-acyl carrier protein synthase III. J. Biol. Chem., 274, 36465-36471 (1999).
- Jones, A.L., Gane, A.M., Herbert, D. et al. β-Ketoacyl-acyl carrier protein synthase III from pea: properties, inhibition by a novel thiolactomycin analogue and isolation of a cDNA encoding the enzyme. Planta, 216, 752-761 (2003).
- Slabas, A.R., Chase, D., Nishida, I. et al. Molecular cloning of higher-plant 3-oxoacyl-(acyl carrier protein) reductase. Sequence identities with the nodG-gene product of the nitrogen-fixing soil bacterium Rhizobium meliloti. Biochem. J., 283, 321-326 (1992).
- Fawcett, T., Copse, C.L., Simon, W.J. and Slabas, A.R. Kinetic mechanism of NADH-enoyl-ACP reductase from Brassica napus. FEBS Lett., 484, 65-68 (2000).
- Levy, C.W., Roujeinikova, A., Sedelnikova, S. et al. Molecular basis of triclosan activity. Nature, 398, 383-384 (1999).
- Browse, J. and Somerville, C. Glycerolipid synthesis: biochemistry and regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42, 467-506 (1991).
- Eccleston, V.S. and Ohlrogge, J.B. Expression of lauroyl-ACP thioesterase in Brassica napus seeds induces pathways for both fatty acid oxidation and biosynthesis and implies a set point for triacylglycerol accumulation. Plant Cell, 10, 613-621 (1998).
- Wallis, J.G. and Browse, J. Mutants of Arabidopsis reveal many roles for membrane lipids. Prog. Lipid Res., 41, 254-278 (2002).
- Shanklin, J. and Cahoon, E.B. Desaturation and related modifications of fatty acids. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49, 611-641 (1998).
- Hildebrand, D.F., Yu, K., McCracken, C. and Rao, S.S. Fatty acid manipulation. In: Plant Lipids: Biology, Utilisation and Manipulation, pp. 67-102 (D.J. Murphy (ed.) Blackwell Publishing, Oxford) (2005).
- Domonkos, I., Laczko-Dobos, H. and Gombos, Z. Lipid-assisted protein-protein interactions that support photosynthetic and other cellular activities. Prog. Lipid Res., 47, 422-435 (2008).
- Murata, N. and Wada, H. Acyl lipid desaturases and their importance in the tolerance and acclimatization to cold of cyanobacteria. Biochem. J., 308, 1-8 (1995).
- Napier, J.A. The production of unusual fatty acids in transgenic plants. Annu. Rev. Plant Biol., 58, 295-319 (2007).
- Dyer, J. and Mullen, R. Development and potential of genetically engineered oilseeds. Seed Sci. Res., 15, 255-267 (2005).
- Jaworski, J. and Cahoon, E.B. Industrial oils from transgenic plants. Curr. Opin. Plant Biol., 6, 178-184 (2003).
- Thelan, J.J. and Ohlrogge, J. Metabolic engineering of fatty acid biosynthesis in plants. Metab. Eng., 4, 12-21 (2002).
- Voelker, T. and Kinney, A.J. Variations in the biosynthesis of seed-storage lipids. Annu. Rev. Plant Phys., 52, 335-361 (2001).
- Cunnane, S.C. Problems with essential fatty acids: time for a new paradigm? Prog. Lipid Res., 42, 544-568 (2003).
- Metz, J.G., Roessler, P., Facciotti, D. et al. Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science, 293, 290-293 (2001).
- von Wettstein-Knowles, P. Biosynthesis and genetics of waxes. In: Waxes: Chemistry, Molecular Biology and Functions, pp. 91-130 (R. J. Hamilton (ed.) The Oily Press, Dundee) (1995).
- Kunst, L., Samuels, A.L. and Jetter, R. The plant cuticle: formation and structure of epidermal surfaces. In: Plant Lipids: Biology, Utilisation and Manipulation, pp. 270-302 (D.J. Murphy (ed.) Blackwell Publishing, Oxford) (2005).
Additional resource
In This Section
- Plant Fatty Acid Synthesis
- Production of Unusual Fatty Acids in Plants
- Arabidopsis Acyl-Coenzyme A-Binding Proteins
- Long Chain acyl-coA Synthetases and Other Acyl Activating Enzymes
- Plant Triacylglycerol Synthesis
- Triacylglycerol Biosynthesis in Eukaryotic Microalgae
- Subcellular Oil Droplets and Oleosins in Plants
- Triacylglycerol Mobilisation in Plants
- Role of Transcription Factors in Storage Lipid Accumulation in Plants
- Biosynthesis of Plant Lipid Polyesters
- Rubber Biosynthesis in Plants
- Carotenoid Biosynthesis and Regulation in Plants
- The Oxylipin Biosynthetic Pathways in Plants
- N-Acylphosphatidylethanolamines (NAPEs), N-acylethanolamines (NAEs) and Other Acylamides: Metabolism, Occurrence and Functions in Plants
- Phosphoinositide Signaling in Plants
- Plant Lipidomics
- 50 years of Galactolipid Research: The Beginnings
- Transport and function of lipids in the plant phloem