Carotenoid Biosynthesis and Regulation in Plants
The Authors: Abdelali Hannoufa and Zakir Hossain, Agriculture and Agri-Food Canada, 1391 Sandford Street, London, Ontario, Canada
Introduction
Carotenoids are lipophilic secondary metabolites derived from the isoprenoid pathway and are accumulated in most plant organs [1]. They contribute to the red, yellow and orange colors of many fruits and flowers, and are a factor in attracting pollinators to flowers. Carotenoids can exert important physiological functions in a wide range of organisms, including plants and humans. They are essential components of the photosynthetic machinery, and play a critical role in preventing photooxidative damage [1]. Carotenoid catabolism products, such as β-ionones, are involved in plant-insect interactions. The importance of carotenoids for plant growth and development is evident since at least two major phytohormones, strigolactones and abscisic acid, are derived from carotenoid precursors [2]. Some carotenoids are precursors of vitamin A, and prevent human age-related macular degeneration. Others, like lycopene, a red carotenoid pigment contained in tomato and watermelon, is a potent antioxidant and is considered to prevent prostate cancer. Astaxanthin, another red carotenoid, is found mainly in red sea animals and some algae and is likely to prevent cardiovascular disease and UV-light aging in the human body. Carotenoids are also widely used as colorants in the food and cosmetic industries, and some are important supplements in livestock and fish feed formulations.
Although carotenoid biosynthesis in plants has been well investigated, extensive studies on its regulation are relatively limited. Knowledge of carotenoid biosynthesis and regulation has led to a plethora of successful attempts at metabolic engineering of carotenoids in economically important crops [3]. Here, we briefly summarize carotenoid biosynthesis, and identify steps that control the flux through the pathway. We also discuss how in addition to the rate of biosynthesis, sequestration and the storage capacity of the cell, along with the rate of carotenoid catabolism, all play a significant role in determining the levels of carotenoid accumulation in plant tissues and organs.
Carotenoid Biosynthesis
Since the landmark review paper by Cunningham and Gantt [4], major discoveries have been made in the characterization of carotenoid biosynthesis and its regulation in plants (for the latest updates, see [5,6]; and Figure 1 for a compressed version of the carotenoid biosynthesis pathway). The core carotenoid pathway is conserved in most plant species although some plants accumulate special and rare carotenoids via unique biosynthetic routes. As isoprenoids, carotenoid compounds originate in the plastid-localized 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway that starts with the reaction between pyruvate and glyceraldehyde-3-phosphate. The first steps in the MEP pathway are regulated by 1-deoxy-D-xylulose-5-phostaphate synthase (DXS) and 1-deoxy-D-xylulose 5-phosphate reductoisomerase (DXR) (Fig. 1). The second key regulatory step is catalyzed by 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase (HDR) eventually leading to the production of isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP) (Fig. 1). Geranyl-geranyl diphosphate (GGPP) synthase catalyses the condensation of three molecules of IPP and one molecule of DMAPP to produce GGPP – a 20-carbon molecule. The first committed step in carotenoid biosynthesis is the condensation of two molecules of GGPP by phytoene synthase (PSY) to form phytoene (Fig. 1). GGPP is also the precursor for several other groups of metabolites, including chlorophylls, ubiquinones and tocopherols. Phytoene then undergoes four sequential reactions to form lycopene.
Figure 1. Carotenoid biosynthesis pathways
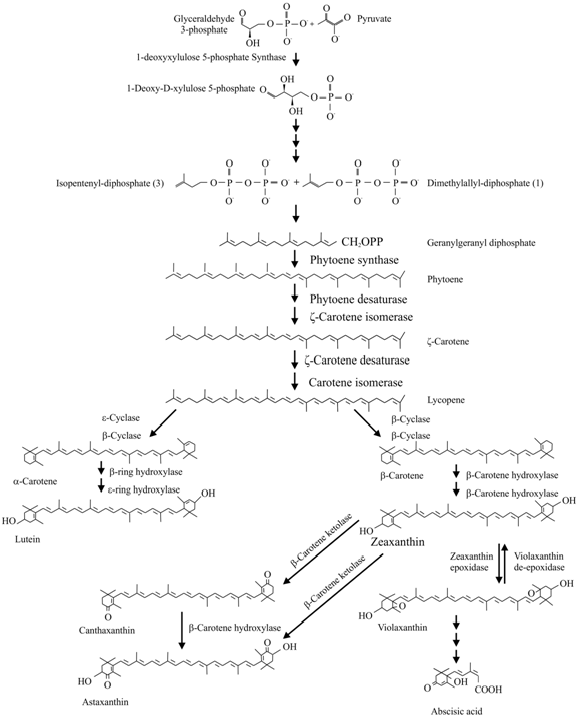
In bacteria, only one phytoene desaturase, crtI, catalyzes the conversion of phytoene to lycopene; however in plants, at least four enzymes are required for this step. These enzymes are phytoene desaturase (PDS) and zeta-carotene desaturase (ZDS) which produce respective poly-cis-compounds, which are then isomerized to trans forms by zeta carotene isomerase (ZISO) and carotenoid isomerase (CRTISO) to produce lycopene.
In higher plants, the cyclization of lycopene with lycopene ε- and β-cyclases is a critical branch-point in carotenoid biosynthesis (Fig. 1) [2]. In one branch, a single enzyme, lycopene β-cyclase (β-CYC), introduces a β-ring at both ends of lycopene to form β-carotene in a sequential two-step reaction. The first dedicated reaction in the other branch, leading to lutein, requires both ε-CYC and lycopene β-cyclase (β-CYC) to introduce one β- and one ε-ring into lycopene to form α-carotene. α-Carotene is acted upon by a β-ring hydroxylase to form zeinoxanthin, which is then hydroxylated by an ε-ring hydroxylase to produce lutein. Unlike plants, the cyanobacterium Prochlorococcus marinus MED4 encodes a ε-CYC that can simultaneously catalyze the formation of α-, β- and ε-carotenes. Carotenoids with two ε-rings are rare in plants and algae, with an exception in lettuce, wherein a single ε-CYC adds two ε-rings to lycopene to form lactucaxanthin [7]. β-Carotene can be hydroxylated in a two-step reaction to zeaxanthin, with β-cryptoxanthin as an intermediate product. In green tissues, zeaxanthin can be epoxidized to violaxanthin, and a set of light- and dark-controlled reactions known as the xanthophyll cycle rapidly optimize the concentration of violaxanthin and zeaxanthin in the cell through the action of zeaxanthin epoxidase and violaxanthin de-epoxidase, respectively, via antheraxanthin [8].
A myriad of other carotenoids with unique biosynthetic routes exist in some plants. For example, some higher plants, such as Adonis aestivalis, and cyanobacteria accumulate the ketocarotenoid astaxanthin, which is derived from zeaxanthin oxidation; a reaction catalysed by β-carotene ketolase. Astaxanthin can also result from canthaxanthin hydroxylation, which is catalyzed by β-carotene hydroxylase. Canthaxanthin biosynthesis from β-carotene is catalyzed by β-carotene ketolase. An alternative pathway for astaxanthin biosynthesis in A. aestivalis was also suggested [9]. The red varieties of pepper accumulate two ketoxanthophylls, capsanthin and capsorubin, which account for the red color of the fruit. A single bifunctional enzyme, capsanthin-capsorubin synthase, was shown to be responsible for converting antheraxanthin to capsanthin, and violaxanthin to capsorubin [10].
Regulation of Carotenoid Biosynthesis
Carotenoid accumulation occurs in most plant tissues, including green shoots, flowers, fruits, seeds and roots. Although the contents and types of carotenoids of green tissues are relatively conserved across most plant species, the levels of carotenoids and their profiles in nongreen tissues, such as flowers, fruits and seeds, vary considerably, and are influenced by many factors, including the developmental stage, environment, stress or a combination of these [1]. In general, the steady-state levels of carotenoids are determined by the rate of biosynthesis, storage capacity of the cell and the rate of catabolism and degradation. Combined, these factors have made the study of carotenoid regulation challenging. Here, we discuss only the regulation of carotenoid biosynthesis as determined by the main ‘bottlenecks’ in the biosynthesis pathway. We also briefly discuss how knowledge of these important steps has been used in the metabolic engineering of carotenoid biosynthesis in plants. The ability of the cell to sequester carotenoids and the rate of carotenoid catabolism will also be discussed as additional factors used to determine the levels of carotenoids.
Isopentenyl pyrophosphate (IPP) biosynthesis
The fact that the first reaction in the MEP pathway is catalyzed by the DXS enzyme makes it a presumptive regulatory step in carotenoid biosynthesis. The initial evidence in support of this came from results of overexpressing and silencing of the DXS gene in Arabidopsis seedlings. Overexpression of DXS was shown to result in up to 112–131% increase in the total carotenoid content, whereas silencing of this gene reduced the carotenoid content by 75–87% relative to the wild-type control. In ripening tomato fruits, the abundance of DXS transcript was found to be developmentally regulated, and corresponded to changes in expression of PSY and accumulation of carotenoids [11]. DXS has been exploited in biotechnological applications to improve the carotenoid content of crops. For example, its overexpression in potato caused a ~2–fold increase in tuber carotenoid content, with a 6─7-fold increase in phytoene content relative to the control [12].
Phytoene biosynthesis
Phytoene biosynthesis is the first committed step in carotenoid biosynthesis, and has long been considered a ‘bottleneck’ in the pathway. Except for Arabidopsis, most other plant species express multiple functionally redundant copies of phytoene synthase (PSY), although different PSY genes appear to be differentially expressed and regulated. At least three PSY homologues are encoded by the maize genome, and the expression of one of them, PSY3, was induced by salt and drought, and this induction resulted in enhanced carotenoid content. Differential regulation of the three members of PSY by light (PSY1 and PSY2) and ABA (PSY3) in rice was reported to correlate with the presence of cis-acting elements responsive to the two conditions in their respective promoters [13]. In Arabidopsis, its single copy PSY gene is tightly regulated by light. A phytochrome-interacting transcription factor, RIF, binds to the PSY promoter and maintains it in a repressed state under dark conditions. Under light conditions, RIF degrades and dissociates from the PSY promoter, thus allowing for its active expression [14]. Another transcription factor, RAP2.2, was also shown to bind to the PSY promoter and regulate PSY expression and carotenoid levels. PSY is also a limiting factor in carotenoid biosynthesis in the case of Brassica napus, and PSY genes from several species have been used to increase carotenoid levels in plants.
Overexpression of a bacterial phytoene synthase (crtB) fused to a plastid transit peptide and the seed-specific napin promoter caused up to a 300-fold increase in β-carotene accumulation in the seeds of B. napus. Carotenoid levels increased in vegetative tissues of transgenic tomato plants, and a concomitant dwarf phenotype was observed in the transformants when the PSY gene was constitutively overexpressed. To overcome this adverse effect, the PSY gene was spatially and temporally expressed which increased the carotenoid content in ripening fruits without such adverse effects. The PSY gene(s) have also been used to increase the carotenoid contents of staple foods, such as rice. Overexpression of the maize PSY gene resulted in higher carotenoid levels in the rice endosperm [15].
Lycopene biosynthesis
The first step in lycopene biosynthesis is catalyzed by phytoene desaturase (PDS). The promoter of the Arabidopsis PDS gene was found to have a regulatory region similar to that of the binding site of the RAP2.2 transcription factor to the PSY promoter, and analysis showed that RAP2.2 does indeed bind to the PDS promoter and affect carotenoid accumulation. Carotenoid isomerization catalyzed by CRTISO, which isomerizes cis bonds to all-trans lycopene, is another step with regulatory impact on carotenoid biosynthesis. Characterization of the ccr1 locus of Arabidopsis revealed that it encodes a histone methyltransferase (SET DOMAIN GROUP, SDG8). This enzyme catalyzes the methylation of chromatin histones, and when the expression of SDG8 gene was disrupted, it caused a reduction in the levels of CRTISO transcript and carotenoid content [16]. In addition, Arabidopsis plants with reduced SDG8 expression showed altered shoot and root branching, which were possibly caused by changes in levels of branch-inhibiting strigolactone hormones that are derived from carotenoid catabolism products [2]. This was the first report on epigenetic regulation of carotenoid biosynthesis; this breakthrough has the potential to open new avenues for engineering of carotenoids in plants.
Lycopene cyclization
The carotenoid biosynthesis pathway branches after the formation of lycopene. One branch forms carotenoids with two β-rings, while the other introduces both β- and ε- rings to lycopene to form α-carotene, which is then converted to lutein. Thus, the relative activities of β-CYC and ε-CYC would be expected to determine the proportion of lycopene channeled to the two branches of the carotenoid pathway; i.e. β,β- and ε,β-carotenoids [16-18]. Flux through the carotenoid pathway may play a role in controlling ε-CYC. For example, Arabidopsis ccr1 (defective in SDG8) and ccr2 (defective CRTISO) had lower lutein content and ε-CYC transcript levels [17]. Manipulation of expression of β-CYC and ε-CYC genes has been exploited to alter the ratios of the β,β- and ε,β-carotenoids in crops. Indeed, transgenic manipulations of ε-CYC and β-CYC expression in Arabidopsis, potato, tomato, maize and B. napus altered the pool of β-ring–containing carotenes and xanthophylls [17-19].
In Arabidopsis, the lut2 mutant is deficient in ε-CYC and its leaves are devoid of lutein, but it has high levels of β-carotene, the product from the competing branch [17]. Silencing of ε-CYC by co-suppression also caused changes in the ratios of lutein to β-carotene in Arabidopsis [17]. Overexpression of β-CYC in tomato plastids resulted in an unexpected increase in the total carotenoid content, with predominant accumulation of β-carotene in the fruits and xanthophyll carotenoids in the leaves [20]. Downregulation of ε-CYC in B. napus using RNAi resulted in seeds with reduced expression of ε-CYC and increased levels of total carotenoids, including β-carotene, zeaxanthin, violaxanthin and, unexpectedly, lutein [19]. In potato, tuber-specific silencing of ε-CYC resulted in increased levels of β-carotene (up to 14–fold), with two transgenic lines showing slight increases in lutein (1.5-1.8 fold). Natural variations in ε-CYC in Zea mays has been used to select for varieties with varying levels of β,β- and ε,β-carotenoids [21]. In their study, Harjes et al. [21], identified four polymorphisms that accounted for 58% of variations in the levels of the two sets of carotenoids. Collectively, these findings highlight the role of lycopene epsilon cyclization as a major regulatory step in carotenoid biosynthesis determining the ratios of β,β- and ε,β-carotenoids in plants.
Sequestration and storage
In addition to the rate of carotenoid biosynthesis, other factors, such as sequestration and availability of storage compartments, play a significant role in determining levels of carotenoid accumulation. Even though carotenoid biosynthetic enzymes are nuclear-encoded, they are all located in the plastids, where carotenoids are synthesized and accumulated. In chloroplasts, most carotenoids accumulate in the form of chlorophyll–carotenoid–protein complexes in the thylakoid membranes associated with light-harvesting antenna. These complexes play an important role in stabilizing plant light–harvesting complexes (LHCs) and in assembling a functional photosystem II (PSII). Seed carotenoids are compartmentalized to elaioplasts (lipid-storing plastids), which use specialized lipoprotein-sequestering structures to store large quantities of carotenoids [1]. In chromoplasts, significant amounts of carotenoids may be stored in membranes, oil bodies or other crystalline structures within the stroma [1,4].
Reports in the literature have shown cases where enhanced carotenoid accumulation was accompanied by changes in the anatomical structure of plastids, resulting in enhanced ability to store carotenoids [22]. For example, a mutation in Brassica oleracea Orange (Or) gene resulted in enhanced carotenoid content in the curds, but no significant change was detected in the expression of the major carotenogenic genes [22]. Further analysis revealed changes in the thylakoid membranes of the plastids, thus allowing them to store large quantities of carotenoids [22]. Overexpression of PSY in B. napus seeds also resulted in increased carotenoid accumulation, and the plastids of transgenic seeds were found to have thread-like inclusion bodies, which were proposed to accumulate the excess carotenoids. In addition, changes in carotenoid accumulation during B. napus seed development, and those resulting from silencing of ε-CYC, did not correspond to changes in the expression of carotenogenic genes [19]. The authors therefore suggested that enhanced carotenoid accumulation could be due to changes in the cell structure, thus allowing for storage of more carotenoids.
Catabolism and degradation
A recent study revealed that carotenoids are constantly synthesized and degraded [23]. This presumably maintains carotenoids at physiological levels, especially in green tissues, where a constant ratio of chlorophylls to carotenoids has to be maintained to ensure the integrity of the photosynthesis system. Carotenoid degradation by enzymatic oxidative cleavage produces an array of terpenoid products collectively known as apocarotenoids. These include abscisic acid and strigolactones, and other volatile and nonvolatile compounds, which are well known in respective industries for their use as aromas, flavors and fragrances. Some apocarotenoids, e.g. β-ionone, are also known to play a role in plant-insect interactions.
In Arabidopsis, the gene family that encodes carotenoid cleavage enzymes is comprised of at least nine members. Four of these encode carotenoid cleavage dioxygenases (CCD), and the remaining five encode 9-cis-expoxycarotenoid dioxygenases (NCED), with many of these enzymes exhibiting substrate promiscuity in vitro [24]. The functions of some of these enzymes have been reported to be associated with certain apocarotenoids in several plant species. For example, CCD7 and CCD8 act in a coordinated manner in strigolactone biosynthesis, CCD1 is involved in β-ionone biosynthesis, and NCED2, NCED3, NCED5, NCED6 and NCED9 are associated with abscisic acid production [5]. In chrysanthemums, CCD activity is important for petal color, and elevated transcript levels of CmCCD4a is reported to play a role in the degradation of yellow carotenoid pigments, which resulted in flowers with white petals.
Conclusion
Considering the importance of carotenoids for industry, human health and plant development, emphasis has been given to targeted manipulation of carotenoid biosynthesis in higher plants. In recent years, the carotenoid biosynthetic pathway in higher plants including Arabidopsis, maize, rice, potato, tomato and canola has successfully been engineered through transgenic approaches. However, due to the complex regulation of the carotenoid biosynthesis, modulation of regulatory elements such as transcription factors is important for visible and long-lasting success of carotenoid metabolic engineering. The recent discovery of novel regulators, such as DET1 and microRNA156b [25], Or [22] and SDG8 [16] will undoubtedly allow for more precise enrichment of crops with carotenoids.
References
- Howitt, C.A. and Pogson, B.J. Carotenoid accumulation and function in seeds and non-green tissues. Plant Cell Environ., 29, 435-445 (2006) (DOI: 10.1111/j.1365-3040.2005.01492.x).
- Cazzonelli, C.I. and Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci., 15, 266-274 (2010) (DOI: 10.1016/j.tplants.2010.02.003).
- Giuliano, G., Tavazza, R., Diretto, G., Beyer, P. and Taylor, M.A. Metabolic engineering of carotenoid biosynthesis in plants. Trends Biotechnol., 26, 139-145 (2008) (DOI: 10.1016/j.tibtech.2007.12.003).
- Cunningham, F.X. and Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49, 557-583 (1998) (DOI: 10.1146/annurev.arplant.49.1.557).
- Walter, M.H. and Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep., 28, 663-692 (2011) (DOI: 10.1039/c0np00036a).
- Farré, G., Sanahuja, G., Naqvi, S., Bai, C., Capell, T., Zhua, C. and Christou, P. Travel advice on the road to carotenoids in plants. Plant Sci., 179, 28-48 (2010) (DOI: 10.1016/j.plantsci.2010.03.009).
- Cunningham, F. and Gantt, E. One ring or two? Determination of ring number in carotenoids by lycopene ɛ-cyclases. Proc. Natl. Acad. Sci. U.S.A., 98, 2905-2910 (2001) (DOI: 10.1073/pnas.051618398).
- Demmig-Adams, B. and Adams, W.W. Antioxidants in photosynthesis and human nutrition. Science, 298, 2149-2153 (2002) (DOI: 10.1126/science.1078002).
- Cunningham, F.X. and Gantt, E. A study in scarlet: Enzymes of ketocarotenoid biosynthesis in the flowers of Adonis aestivalis. Plant J., 41, 478-492 (2005) (DOI: 10.1111/j.1365-313X.2004.02309.x).
- Bouvier, F., Hugueney, P., D'Harlingue, A., Kuntz, M. and Camara, B. Xanthophyll biosynthesis in chromoplasts: Isolation and molecular cloning of an enzyme catalyzing the conversion of 5,6-epoxycarotenoid into ketocarotenoid. Plant J., 6, 45-54 (1994) (DOI: 10.1046/j.1365-313X.1994.6010045.x).
- Lois, L.M., Rodríguez-Concepción, M., Gallego, F., Campos, N. and Boronat, A. Carotenoid biosynthesis during tomato fruit development: Regulatory role of 1-deoxy-D-xylulose 5-phosphate synthase. Plant J., 22, 503-513 (2000) (DOI: 10.1046/j.1365-313X.2000.00764.x).
- Morris, W.L., Ducreux, L.J.M., Hedden, P., Millam, S. and Taylor, M.A. Overexpression of a bacterial 1-deoxy-D-xylulose 5-phosphate synthase gene in potato tubers perturbs the isoprenoid metabolic network: Implications for the control of the tuber life cycle. J. Exp. Bot., 57, 3007-3018 (2006) (DOI: 10.1093/jxb/erl061).
- Welsch, R., Wüst, F., Bär, C., Al-Babili, S. and Beyer, P. A third phytoene synthase is devoted to abiotic stress-induced abscisic acid formation in rice and defines functional diversification of phytoene synthase genes. Plant Physiol., 147, 367-380 (2008) (DOI: 10.1104/pp.108.117028).
- Toledo-Ortiz, G., Huq, E. and Rodríguez-Concepción, M. Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proc. Natl. Acad. Sci. U.S.A., 107, 11626-11631 (2010) (DOI: 10.1073/pnas.0914428107).
- Paine, J.A., Shipton, C.A., Chaggar, S., Howells, R.M., Kennedy, M.J., et al. Improving the nutritional value of golden rice through increased pro-vitamin A content. Nat. Biotechnol., 23, 482-487 (2005) (DOI: 10.1038/nbt1082).
- Cazzonelli, C.I., Roberts, A.C., Carmody, M.E. and Pogson, B.J. Transcriptional control of SET DOMAIN GROUP 8 and CAROTENOID ISOMERASE during Arabidopsis development. Mol. Plant, 3, 174-191 (2010) (DOI: 10.1093/mp/ssp092).
- Pogson, B., McDonald, K.A., Truong, M., Britton, G. and DellaPenna, D. Arabidopsis carotenoid mutants demonstrate that lutein is not essential for photosynthesis in higher plants. Plant Cell, 8, 1627-1639 (1996) (DOI: 10.1105/tpc.8.9.1627).
- Cunningham, F.X., Pogson, B., Sun, Z., McDonald, K.A., DellaPenna, D., et al. Functional analysis of the β and ε lycopene cyclase enzymes of Arabidopsis reveals a mechanism for control of cyclic carotenoid formation. Plant Cell, 8, 1613-1626 (1996) (DOI: 10.1105/tpc.8.9.1613).
- Yu, B., Lydiate, D.J., Young, L.W., Schäfer, U.A. and Hannoufa, A. Enhancing the carotenoid content of Brassica napus seeds by downregulating lycopene epsilon cyclase. Transgenic Res., 17, 573-585 (2008) (DOI: 10.1007/s11248-007-9131-x).
- Apel, W. and Bock, R. Enhancement of carotenoid biosynthesis in transplastomic tomatoes by induced lycopene-to-provitamin A conversion. Plant Physiol., 151, 59-66 (2009) (DOI: 10.1104/pp.109.140533).
- Harjes, C.E., Rocheford, T.R., Bai, L., Brutnell, T.P., Kandianis, C.B., et al. Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification. Science, 319, 330-333 (2008) (DOI: 10.1126/science.1150255).
- Paolillo, D.J., Garvin, D.F. and Parthasarathy, M.V. The chromoplasts of Or mutants of cauliflower (Brassica oleracea L. var. botrytis). Protoplasma, 224, 245-253 (2004) (DOI: 10.1007/s00709-004-0059-1).
- Beisel, K.G., Jahnke, S., Hofmann, D., Köppchen, S., Schurr, U., et al. Continuous turnover of carotenes and chlorophyll a in mature leaves of Arabidopsis revealed by 14CO2 pulse-chase labeling. Plant Physiol., 152, 2188-2199 (2010) (DOI: 10.1104/pp.109.151647).
- Vogel, J.T., Walter, M.H., Giavalisco, P., Lytovchenko, A., Kohlen, W., et al. Slccd7 controls strigolactone biosynthesis, shoot branching and mycorrhiza-induced apocarotenoid formation in tomato. Plant J., 61, 300-311 (2010) (DOI: 10.1111/j.1365-313X.2009.04056.x).
- Wei, S., Yu, B., Gruber, M.Y., Khachatourians, G.G., Hegedus, D.D. and Hannoufa, A. Enhanced seed carotenoid levels and branching in transgenic Brassica napus expressing the Arabidopsis miR156b gene. J. Agric. Food. Chem., 58, 9572-9578 (2010) (DOI: 10.1021/jf102635f).
In This Section
- Plant Fatty Acid Synthesis
- Production of Unusual Fatty Acids in Plants
- Arabidopsis Acyl-Coenzyme A-Binding Proteins
- Long Chain acyl-coA Synthetases and Other Acyl Activating Enzymes
- Plant Triacylglycerol Synthesis
- Triacylglycerol Biosynthesis in Eukaryotic Microalgae
- Subcellular Oil Droplets and Oleosins in Plants
- Triacylglycerol Mobilisation in Plants
- Role of Transcription Factors in Storage Lipid Accumulation in Plants
- Biosynthesis of Plant Lipid Polyesters
- Rubber Biosynthesis in Plants
- Carotenoid Biosynthesis and Regulation in Plants
- The Oxylipin Biosynthetic Pathways in Plants
- N-Acylphosphatidylethanolamines (NAPEs), N-acylethanolamines (NAEs) and Other Acylamides: Metabolism, Occurrence and Functions in Plants
- Phosphoinositide Signaling in Plants
- Plant Lipidomics
- 50 years of Galactolipid Research: The Beginnings
- Transport and function of lipids in the plant phloem